 2024, Vol. 55
2024, Vol. 55中国海洋湖沼学会主办。
文章信息
- 韦毓, 翁旭东, 于瑾, 高阳, 应晓国, 余林, 胡家杰, 曾妮. 2024.
- WEI Yu, WENG Xu-Dong, YU Jin, GAO Yang, YING Xiao-Guo, YU Lin, HU Jia-Jie, ZENG Ni. 2024.
- 不同养殖模式对鲫鱼(Carassius auratus)营养品质及特征风味的影响研究
- STUDY ON THE EFFECTS OF DIFFERENT CULTURE MODES ON THE NUTRITIONAL QUALITY AND CHARACTERISTIC FLAVOR OF CARASSIUS AURATUS
- 海洋与湖沼, 55(1): 243-252
- Oceanologia et Limnologia Sinica, 55(1): 243-252.
- http://dx.doi.org/10.11693/hyhz20230800171
文章历史
-
收稿日期:2023-08-23
收修改稿日期:2023-11-26




2. 浙江渔老大农业科技有限公司 浙江衢州 324400;
3. 龙游县农业农村局养殖业发展中心 浙江衢州 324400
2. Zhejiang Yulaoda Agricultural Technology Co., Ltd., Quzhou 324400, China;
3. Longyou Aquaculture Development Center, Agricultural and Rural Bureau of Longyou County, Quzhou 324400, China
我国是水产养殖大国, 其中淡水鱼产量在全球占主导地位(蔡振鑫等, 2021)。鲫鱼(Carassius auratus)作为我国重要养殖的经济淡水鱼种, 其因鲜美细嫩的肉质和丰富的营养价值而深受消费者喜爱。鲫鱼的养殖方式通常为传统池塘养殖, 即“三塘合一”的生态系统模式, 该模式下鱼类生存环境、天然饵料生产基地、有机物分解场所均为一处, 因而常出现水体富营养化、养殖系统失衡等问题(刘凯等, 2021)。近年来, 池塘内循环流水跑道养殖模式逐步兴起, 跑道养殖是一种将池塘分为推水区、养殖区和净化区的高效养殖模式, 其中推水区主要利用机械装置使养殖水体处于循环流水状态(贾丽等, 2011; 李倩等, 2020)。同时相较于传统池塘养殖, 具有资源节约、循环生态的优势, 还能使养殖对象较高效的吸收利用氮磷(刘梅等, 2021), 该模式有望解决传统池塘养殖所存在的部分问题。
不同养殖方式对鱼类营养品质和风味会产生一定影响。根据研究报道(朱士臣等, 2021; 黄爱霞等, 2021; 李敏等, 2022)可知, 跑道养殖模式可改善青鱼、梭鱼、三角鲂等的营养品质和风味口感, 从而提高了消费者的可接受度和鱼类价值。刘梅等(2019)的研究指出, 通过调节水流速度可以改变鱼的运动状态、影响脂肪沉积等指标, 而跑道养殖模式对黄颡鱼生长性能、形体指标、血清生化指标及肌肉营养成分均具有改善作用。目前, 关于跑道养殖和池塘养殖模式对鲫鱼营养品质和特征风味影响研究较少。
因此, 本研究对跑道养殖和传统池塘养殖模式下的鲫鱼形体、基础营养成分、蒸煮损失和持水力、氨基酸、脂肪酸、肌肉蛋白质消化率和挥发性化合物等指标进行研究比较, 旨在探究养殖方式对鲫鱼营养品质及特征风味的影响, 以期为选择适宜养殖模式、提高鲫鱼质量及快速鉴别鲫鱼的养殖模式提供理论参考。
1 材料与方法 1.1 材料与试剂实验材料: 鲫鱼购自衢州龙游龙和渔业养殖基地(119°178339′E, 29°000213′N)。鲫鱼来自同一批次健康幼鱼, 分别投入两组池塘(传统养殖池塘和内循环流水养殖系统池塘), 期间投喂统一配方的饲料, 养殖6个月后, 随机从两种养殖模式中取体型完整、灵敏健康的鲫鱼各6尾, 活体运送至实验室, 分别命名为跑道组[体重(390.32±76.02) g, 体长(29.08±1.91) cm]和池塘组[体重(282.68±27.55) g, 体长(25.22±0.84) cm]。
实验试剂: 双缩脲试剂盒, 南京建成生物工程研究所; 胃蛋白酶(猪胃黏膜), 上海瑞永生物科技有限公司; 胰蛋白酶(猪胰), 上海瑞永生物科技有限公司; 其他试剂均为分析纯, 国药集团化学试剂公司。
1.2 仪器与设备脂肪测定仪SZT-06A, 苏州市天威仪器有限公司; 恒温振荡器THZ-98A, 上海一恒科学仪器有限公司; 高速离心机Multifuge X1R, 美国ThermoFisher Scientific公司; 日立L-8900型氨基酸全自动分析仪, 日本日立高新技术公司; Agilent6890型气相色谱仪, 美国安捷伦科技公司; FlavourSpec® GC-IMS, 德国G.A.S公司。
1.3 实验方法 1.3.1 样品处理分别测量两者鱼体长度和质量, 随后解剖鱼体取出内脏(主要为肝脏)并称重, 以计算脏体比, 肝体比和肥满度。最后, 鱼胴体洗净后置于4 ℃冰箱备用。
1.3.2 基础营养成分含量测定水分含量的测定采用《GB 5009.3-2016》中的直接干燥法进行; 粗脂肪含量的测定采用《GB 5009.6-2016》中的索氏抽提法进行; 粗灰分含量的测定采用《GB 5009.4-2016》中的马弗炉高温灼烧法进行。总蛋白含量测定采用南京建成生物工程研究所双缩脲试剂盒。
1.3.3 蒸煮损失和持水力蒸煮损失: 将鱼肉斩拌后准确称取质量, 记为W1, 放入蒸煮袋后密封, 将其置于90 ℃恒温水浴中加热10 min, 取出放置恢复至室温后, 用滤纸吸干样品表面水分称重, 记为W2。蒸煮损失按公式(1)计算:
 (1)
(1)持水力: 将鱼肉斩拌后准确称取质量, 记为W3, 将其用三层滤纸包裹后放入50 mL干燥离心管中, 使样品在4 ℃下5 000×g离心15 min, 离心结束后, 取出样品称重, 记为W4。持水力测定按公式(2)计算:
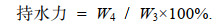 (2)
(2)胃消化率: 测定方法参照孙立春等(2012)等报道并作适当修改。鱼肉被切碎混匀后, 称取3 g样品(记为m0)于锥形瓶中, 加入20 mL的模拟胃液, 将样品置于37 ℃恒温振荡摇床上100 r/min反应4 h, 模拟胃蠕动, 反应结束后, 加入1 mL Na2CO3 (1 mol/L)溶液终止酶反应。消化液在105 ℃烘箱中烘干至恒重, 称量干质量(m1)。体外胃蛋白酶消化率按公式(3)计算:
 (3)
(3)肠消化率: 测定参照孙立春等(2012)的报道并作适当修改。鱼肉切碎混匀后, 称取3 g样品(记为m2)于锥形瓶中, 加入20 mL模拟肠液, 将样品置于37 ℃恒温振荡摇床上100 r/min反应4 h, 模拟肠道蠕动反应结束后, 加入1 mL 1 mol/L Na2CO3溶液终止酶反应。消化液在105 ℃恒温干燥箱中烘干至恒重, 称量干质量(m3)。体外胰蛋白酶消化率按公式(4)计算:
 (4)
(4)采用氨基酸自动分析仪测定氨基酸含量(GB 5009.124-2016); 采用气相色谱仪测定脂肪酸含量(GB 5009.168-2016)。
1.3.6 GC-IMS分析参数样品预处理方法: 取3 g鱼肉样品, 置于20 mL顶空进样瓶中, 45 ℃保持20 min后进样, 每个样品重复3次。
GC-IMS分析方法: 通过FlavourSpec®风味分析仪测定样品挥发性组分。MXT-WAX色谱柱(30 m× 0.53 mm×1 μm), 柱温60 ℃, IMS检测温度为45 ℃, 分析时间35 min, 载气/漂移气为纯度≥99.999%的氮气。漂移气流量为150 mL/min, 载气初始流量为2.0 mL/min保持2 min, 在10 min流速线性增加至10 mL/min, 最后在35 min内线性增加至100 mL/min。GC-IMS分析获得风味成分的峰强度通过Origin软件采用Euclidean最短距离进行聚类分析。
1.4 实验数据处理及分析采用Excel整理实验数据, Origin绘图, SPSS 27进行数据分析, 层次聚类分析使用MetaboAnalyst 5.0 (https://www.metaboanalyst.ca/)进行。实验结果以平均值±标准差表示。使用t检验来评估变量之间的差异, P < 0.05表示差异性显著。
2 结果与分析 2.1 形态学指标不同养殖模式下鲫鱼各项形态学指标如表 1所示。鱼的形态指标在一定程度上能够反映鱼类的感官特性和营养状态。其中脏体比可体现出鱼体内可食用部分的多少, 肝体比可体现出鱼体的健康程度, 肥满度可体现出鱼类的肥瘦程度和生长情况(史庆超等, 2021), 且以上指标皆与鱼体内脂肪的积累有关。两组鲫鱼样品中脏体比与肥满度虽差异不显著(P > 0.05), 但跑道组都略低于池塘组, 且跑道组肝体比显著(P < 0.05)低于池塘组肝体比。因此结果表明跑道组鲫鱼体内脂肪含量较少、可食用部分比例更低, 其原因可能是在跑道养殖模式中所采用的气提式增氧推水装置可使鱼长期处于流水富氧的生长环境中, 运动量较池塘养殖鲫鱼更大。
| 模式 | 脏体比/% | 肝体比/% | 肥满度 |
| 跑道组 | 13±1a | 3±0.5a | 0.016±0.002a |
| 池塘组 | 14±1a | 5±1b | 0.018±0.001a |
| 注: 同一列标注不同字母表示差异性显著(P < 0.05) | |||
不同养殖模式下鲫鱼中基础营养成分含量如表 2所示。池塘组和跑道组的鲫鱼水分和灰分含量差异显著(P < 0.05)。跑道组粗脂肪含量明显低于池塘组(P < 0.05), 而蛋白含量明显高于池塘组(P < 0.05)。这可能由于在池塘内循环养殖的水流作用下, 鲫鱼需要消耗更多的能量来应对运动量的增加, 因此机体的脂肪被分解利用, 从而脂肪含量降低(董立学等, 2021)。有研究报道(Zhang et al, 2021), 运动训练可促进蛋白质在肌肉中的沉积, 跑道组鲫鱼蛋白含量高于池塘组也印证了这一点。由此可知, 跑道模式养殖鲫鱼更符合人们对鱼肉高蛋白低脂肪健康食品的追求。
| 模式 | 水分/% | 灰分/% | 粗脂肪/% | 总蛋白/% |
| 跑道组 | 76.33±0.21b | 0.94±0.01a | 6.47±0.11b | 13.57±0.52a |
| 池塘组 | 77.35±0.17a | 0.83±0.02b | 9.13±0.08a | 11.57±0.74b |
| 注: 同一列标注不同字母表示差异性显著(P < 0.05) | ||||
通过蒸煮损失和持水力的测定, 可以反映该鱼肉的保水性能。如图 1所示, 两组鲫鱼在蒸煮测试中质量都有所损失, 其原因是鱼肉蛋白质的热变性, 导致肌纤维的收缩和内部水分向外迁移(Vaskoska et al, 2021)。此外, 有研究表明运动可以增加肌纤维的密度和直径从而减少离心失水率(Cai et al, 2023), 两组鲫鱼持水力变化符合这一说法。以上表明两种养殖模式下鲫鱼的蒸煮损失、持水力变化较为接近。
 |
| 图 1 蒸煮损失和持水力比较 Fig. 1 Comparison of cooking loss and water holding capacity 注: 相同字母表示无显著差异(P > 0.05) |
如图 2所示, 跑道组鲫鱼肌肉模拟胃消化率为73.92%、肠消化率65.57%, 显著高于池塘组肌肉模拟胃消化率69.77%、肠消化率61.85% (P < 0.05)。食物中的蛋白质先在胃中被胃蛋白酶初步水解, 然后进入小肠, 在胰蛋白酶的作用下进一步被水解为小分子肽及游离氨基酸, 最终被肠壁吸收。有研究(Ding et al, 2022)表明, 脂肪通过小肠的机械蠕动能被乳化成乳状液的形式从而被吸收, 该类乳状液的结构主要由其脂肪相、界面和水相的性质所决定, 当脂肪含量过高时, 部分蛋白质被吸附在油水界面上, 会进一步降低蛋白质的分散程度而影响吸收。本研究中, 跑道组鲫鱼的脂肪含量低于池塘组, 在消化过程中其蛋白质可能较少被吸附, 更大程度地被酶水解, 进而促进消化吸收, 因此跑道组鲫鱼蛋白质消化率呈现出高于池塘组的趋势。
 |
| 图 2 肌肉蛋白质消化特性比较 Fig. 2 Comparison of muscle protein digestion characteristics 注: 不同字母表示有显著差异(P < 0.05) |
由表 3可知, 两种养殖模式的鲫鱼肌肉中共检测到了13种游离氨基酸, 其中池塘组未检出游离脯氨酸。游离氨基酸对肉类食品的风味和品质有重要影响, 目前认为影响鱼肉鲜美的氨基酸主要为天冬氨酸、甘氨酸、谷氨酸、丙氨酸和组氨酸等, 它们的含量决定了对风味的影响程度(李双琦等, 2022)。本实验中跑道组的大部分游离氨基酸含量高于池塘组, 其中鲜味氨基酸(天冬氨酸和谷氨酸)和甜味氨基酸(丙氨酸和谷氨酸)等呈味氨基酸含量显著高于池塘组(P < 0.05)。两组鲫鱼的大部分苦味氨基酸(亮氨酸、异亮氨酸、缬氨酸和组氨酸)含量无显著差异(P > 0.05), 但池塘组鲫鱼赖氨酸含量显著高于跑道组鲫鱼(P < 0.05), 而跑道组鲫鱼精氨酸含量显著高于池塘组鲫鱼(P < 0.05), 因此两组鲫鱼整体所呈现的氨基酸苦味差距较小。总体而言, 跑道模式养殖鲫鱼呈现鲜甜味的氨基酸含量更显著, 故相比池塘养殖鲫鱼可能具有更好地风味。
| 氨基酸名称 | 跑道组质量分数/(g/kg) | 池塘组质量分数/(g/kg) |
| 天冬氨酸 Asp▲ | 0.11±0.02a | 0.032±0.006b |
| 谷氨酸 Glu▲ | 0.2±0.04a | 0.053±0.01b |
| 苏氨酸 Thr△ | 0.16±0.01a | 0.079±0.01b |
| 脯氨酸 Pro△ | 0.014±0.002a | NDb |
| 丙氨酸 Ala△ | 0.2±0.02a | 0.14±0.03b |
| 甘氨酸 Gly△ | 0.26±0.02b | 0.55±0.02a |
| 丝氨酸 Ser△ | 0.11±0.03a | 0.054±0.01b |
| 亮氨酸 Leu● | 0.13±0.02a | 0.12±0.03a |
| 异亮氨酸 IIe● | 0.03±0.01a | 0.016±0.01a |
| 缬氨酸 Val● | 0.073±0.01a | 0.052±0.01a |
| 赖氨酸 Lys● | 0.21±0.04b | 0.37±0.03a |
| 组氨酸 His● | 2.1±0.3a | 1.5±0.3a |
| 精氨酸 Arg● | 0.17±0.02a | 0.096±0.01b |
| 注: ▲表示鲜味氨基酸; △表示甜味氨基酸; ●表示苦味氨基酸; ND表示未检出。同一行字母不同表示差异性显著(P < 0.05) | ||
由表 4可知, 跑道组鲫鱼和池塘组鲫鱼均检出了17种氨基酸, 其中必需氨基酸7种, 半必需氨基酸5种, 非必需氨基酸5种。两组鲫鱼肌肉所含的氨基酸在组成上相对平衡, 含量无显著差异(P > 0.05)。在必需氨基酸中亮氨酸和赖氨酸的含量相对较高, 半必需氨基酸中甘氨酸和精氨酸的含量相对较高, 此外非必需氨基酸中天冬氨酸和谷氨酸的含量相对较高。根据FAO/WHO的理想蛋白质模式, 组成氨基酸的必需氨基酸/总氨基酸(ΣEAA/ΣTAA)为40%左右, 必需氨基酸/非必需氨基酸(ΣEAA/ΣNEAA)为60%以上, 表明蛋白质质量较佳(赵亭亭等, 2018), 跑道组和池塘组鲫鱼ΣEAA/ΣTAA分别为40%、42%, 符合以上指标要求; ΣEAA/ΣNEAA分别为66%、72%, 两者都远远高于60%。综上所述, 两组鲫鱼肌肉均含有丰富的氨基酸, 且氨基酸具有良好的平衡性, 营养价值较高, 均是人体补充氨基酸的优质食物来源和选择。
| 氨基酸名称 | 跑道组质量分数/(g/100g) | 池塘组质量分数/(g/100g) |
| 天冬氨酸 Asp | 1.63±0.05a | 1.56±0.06a |
| 苏氨酸 Thr△ | 0.72±0.04a | 0.70±0.04a |
| 丝氨酸 Ser□ | 0.63±0.05a | 0.59±0.04a |
| 谷氨酸 Glu | 2.35±0.42a | 2.22±0.30a |
| 甘氨酸 Gly□ | 0.79±0.02a | 0.71±0.04b |
| 丙氨酸 Ala | 0.94±0.05a | 0.92±0.03a |
| 胱氨酸 Cys□ | 0.10±0.04a | 0.093±0.03a |
| 缬氨酸 Val△ | 0.78±0.06a | 0.77±0.05a |
| 蛋氨酸 Met△ | 0.37±0.04a | 0.30±0.06a |
| 异亮氨酸 IIe△ | 0.68±0.03a | 0.68±0.02a |
| 亮氨酸 Leu△ | 1.34±0.04a | 1.30±0.03a |
| 酪氨酸 Tyr□ | 0.51±0.05a | 0.47±0.05a |
| 苯丙氨酸 Phe△ | 0.73±0.03a | 0.72±0.02a |
| 赖氨酸 Lys△ | 1.17±0.08b | 1.44±0.1a |
| 组氨酸 His | 0.47±0.02a | 0.42±0.03a |
| 精氨酸 Arg□ | 0.88±0.03a | 0.81±0.04a |
| 脯氨酸 Pro | 0.48±0.04a | 0.42±0.03a |
| 必需氨基酸 ΣEAA | 5.79±0.32a | 5.91±0.32a |
| 总氨基酸 ΣTAA | 14.57±1.09a | 14.12±0.97a |
| 非必需氨基酸 ΣNEAA | 8.78±0.77a | 8.21±0.65a |
| ΣEAA/ΣNEAA | 0.66 | 0.72 |
| ΣEAA/ΣTAA | 0.40 | 0.42 |
| 注: △表示必需氨基酸; □半必需氨基酸。同行字母不同表示差异性显著(P < 0.05) | ||
两种养殖模式的鲫鱼脂肪酸组成, 如表 5所示。脂肪酸对机体健康发挥着重要作用, 跑道组和池塘组鲫鱼分别检出了17和15种脂肪酸。相比总脂肪酸含量, 跑道组(6.145 8 g/100 g)显著高于池塘组(2.556 6 g/100 g), 表明跑道组鲫鱼可以为人体提供更多的脂肪酸。
| 脂肪酸 | 跑道组/(g/100g) | 池塘组/(g/100g) |
| C14:0 肉豆蔻酸◇ | 0.039 0±0.002a | 0.012 8±0.001b |
| C15:0 十五烷酸◇ | 0.006 1±0.0001a | NDb |
| C16:0 十六烷酸◇ | 1.021 0±0.002a | 0.386±0.004b |
| C16:1 棕榈油酸◆ | 0.185 6±0.000 2a | 0.087 5±0.000 3b |
| C18:0 硬脂酸◇ | 0.276 1±0.000 1a | 0.089 3±0.000 1b |
| C18:1n9c 油酸◆ | 2.800 1±0.000 1a | 0.748 2±0.000 2b |
| C18:2n6c 亚油酸◆ | 1.335 9±0.000 3a | 0.350 3±0.000 2b |
| C18:3n6 γ-亚油酸◆ | 0.037 3±0.000 2a | 0.007 1±0.000 1b |
| C18:3n3 α-亚油酸◆ | 0.110 7±0.000 3a | 0.027 7±0.000 6b |
| C20:1 花生烯酸◆ | 0.131 4±0.000 4a | 0.050 7±0.000 3b |
| C20:2 顺-11, 14-二十二碳二烯酸◆ | 0.032 2±0.000 2a | 0.013 2±0.000 2a |
| C20:3n6 顺-8, 11, 14-二十二碳三烯酸◆ | 0.066 8±0.000 3a | 0.029 2±0.000 2b |
| C20:3n3 顺-11, 14, 17-二十碳三烯酸◆ | 0.004 7±0.000 2a | NDb |
| C20:4n6 花生四烯酸◆ | 0.059 2±0.000 8a | 0.045 3±0.005b |
| C22:1n9 芥酸◆ | 0.004 7±0.000 2b | 0.006 4±0.000 1a |
| C20:5n3 顺-5, 8, 11, 14, 17-二十碳五烯酸◆EPA | 0.004 2±0.000 1a | 0.003 6±0.000 1b |
| C22:6n3 顺-4, 7, 10, 13, 16, 19-二十二碳六烯酸◆DHA | 0.031 0±0.000 3b | 0.041 9±0.000 2a |
| 不饱和脂肪酸ΣUFA | 4.803 8 | 1.411 1 |
| 饱和脂肪酸ΣSFA | 1.342 0 | 0.488 5 |
| Σ(EPA+DHA) | 0.035 2 | 0.045 5 |
| 不饱和脂肪酸/饱和脂肪酸(ΣUFA/ΣSFA) | 3.58 | 2.88 |
| 总脂肪酸ΣFA | 6.145 8 | 2.556 6 |
| 注: ◇表示饱和脂肪酸; ◆表示不饱和脂肪酸。同行字母不同表示差异性显著(P < 0.05) | ||
脂肪酸可分为饱和脂肪酸和不饱和脂肪酸, 其中跑道组的饱和脂肪酸值(1.342 0 g/100 g)、不饱和脂肪酸(4.803 8 g/100 g)均高于池塘组的0.488 5 g/100 g和1.411 1 g/100 g。因此跑道组中含量较丰富的饱和脂肪酸包括C18:0硬脂酸, 特别是不饱和脂肪酸中的C16︰1棕榈油酸、C18:2n6c亚油酸以及C18︰1n9c油酸。有报道表明, 硬脂酸可以使得饭后的血脂浓度降低(王珂欣等, 2023)。棕榈油酸能够降低糖异生作用(姜晓凡, 2021), 缓解胰岛素抵抗(高山等, 2020), 有利于血糖的调节, 对糖尿病的治疗具有积极影响。亚油酸有助于降低血液中的胆固醇浓度(Fritsche, 2015)。而油酸具有降低心血管疾病的发生率, 减少炎症等作用(Martins et al, 2023)。此外, 二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)因其保健作用而备受关注, 例如改善脑部功能、缓解神经退行性疾病、抗肿瘤活性以及调节脂质代谢等功能(Zhang et al, 2019)。两组鲫鱼都具有较高的EPA、DHA含量。总体而言, 跑道组鲫鱼的脂肪酸组成更合理均衡, 营养价值相对较高, 可作为人体的优质脂肪来源。据报道, 构成食品特征风味的挥发性成分的生成, 与脂肪酸氧化和脂质分解作用有较大关系。如小分子化合物醛、醇和酮, 由长链不饱和脂肪酸的进一步氧化和分解而产生的, 从而导致鱼的风味和异味(Zhao et al, 2022)。同时脂肪酸可以与美拉德反应化合物反应生成气味阈值较低的风味物质, 因而对风味产生更大的影响(Aaslyng et al, 2017), 本实验中两组鲫鱼的风味成分分析还需进一步讨论。
2.7 不同养殖模式下鲫鱼的GC-IMS分析气相色谱-离子迁移谱联用(GC-IMS)技术是一种新的气相分离检测技术, 具有分析迅速、灵敏度高、进样可变、无需前处理等优点(Wang et al, 2020)。Duan等(2021)利用GC-IMS、电子鼻和电子舌技术, 结合游离氨基酸分析, 对不同地理来源和不同物种的鲑鱼进行了风味成分的分析, 结果证明GC-IMS为4种方法之中确定鲑鱼来源的最佳技术。本研究通过GC-IMS技术用于区分不同养殖模式下鲫鱼的特征风味差异。如图 3a所示, 在横坐标1.0处展现了经过归一化处理的谱图反应离子峰。每个谱图点代表一种挥发性有机物, 蓝色色调表示低浓度, 红色则表示高浓度。通过参照谱图中的信号峰, 形成不同样品之间的差异谱图。将跑道组鲫鱼样品作为参照, 与之对比池塘组样品, 挥发性物质含量的差异明显。红色色调加深表示对应物质浓度较跑道组更高, 而蓝色色调加深则表示浓度更低。这结果表明, 在池塘养殖模式下, 鲫鱼样品与参照样品(跑道组)存在显著差异, 从而验证了不同养殖模式的鲫鱼中存在着挥发性风味物质的差异。该差异可能归因于两种养殖模式下鱼类生长环境和饲养方式的不同, 进一步影响了鱼体内的代谢产物和挥发性有机物的生成。
 |
| 图 3 不同养殖模式下鲫鱼的GC-IMS差异谱图(a)和指纹图谱(b) Fig. 3 GC-IMS difference spectrum (a) and fingerprint (b) of C. auratus from different aquaculture modes |
为深入比较不同养殖模式下鲫鱼挥发性化合物的差异, 研究利用Gallery Plot插件绘制了GC-IMS指纹图谱, 如图 3b。在这两种养殖模式下的鲫鱼中, 共检测到57种挥发性化合物(包括11种单体和11种二聚体)。这些化合物可归类为6种酯类、16种醇类、15种醛类、10种酮类、3种酸类、1种烃类以及6种含硫、含氮、含氧及杂环化合物, 其中以酯类、高级醇和醛酮类化合物为主要成分, 且大多数醛类化合物阈值较低, 对整体风味有较大贡献, 其中酮类和醇类物质阈值较醛类高, 主要表现出水果味、奶油味等令人愉悦的香气(徐永霞等, 2021), 但会有部分酮类化合物可能与其他风味物质相互作用, 使鱼腥味增强(吴俊杰等, 2023)。在跑道组鲫鱼中, 高含量的化合物包括乙偶姻、2-戊酮、丙酮、丙酸乙酯、乙缩醛、3-甲基丁醛、2-甲基丙醛、乙醛、吡啶、2, 3-二甲基吡嗪、2, 5-二甲基噻吩、二甲基硫醚、甲硫基丙醛和乙酸异戊酯等。另一方面, 池塘组鲫鱼中高含量的化合物有2-甲基丙酸、丙酸、乙酸、E-2-壬烯醛、E-2-庚烯醛、E-2-戊烯醛、壬醛、辛醛、庚醛、己醛、丙醛、1-辛烯-3-醇、1-己醇、1-戊醇、3-甲基-1-丁醇、1-戊烯-3-醇、1-丁醇、2-甲基-1-丙醇、1-丙醇、乙醇、2-丙醇、2-丁醇、3-辛酮、2-庚酮、4-甲基-3-戊烯-2-酮、2-丁酮、庚酸乙酯、乙酸乙酯、乙酸甲酯、2-戊基呋喃、3-蒈烯和三甲胺等。池塘组中酮类物质含量高, 可能会有鱼腥味加重的风险, 尤其是还含有1-辛烯-3-醇, 其为低阈值化合物, 具有蘑菇味和鱼腥味(孙慧娟等, 2023)。而三甲胺则在鱼死后的氧化还原反应中形成, 呈现典型的鱼腥味, 是腐败鱼的主要成分之一, 且受到消费者排斥(Heising et al, 2014)。因此, 跑道组鲫鱼中的1-辛烯-3-醇和三甲胺含量较低, 这表明跑道组鲫鱼可能具有更佳的风味质量, 这种差异可能归因于其特定的养殖环境和饲养模式。
2.8 不同养殖模式下鲫鱼的层次聚类分析通过对不同养殖模式下鲫鱼的基础营养成分、游离氨基酸、脂肪酸含量和GC-IMS输出峰强度等数据进行归一化处理, 并对结果进行层次聚类分析, 结果仅呈现了T-test差异显著性前60的指标, 具体见图 4。结果显示, 跑道组(PD)和池塘组(CT)样本之间的聚类距离远大于平行样本内部的距离, 表明不同养殖模式下鲫鱼之间存在显著差异。在基础营养成分方面, 跑道组表现出相对较高的蛋白质和灰分含量, 同时呈现较低的粗脂肪含量。然而, 跑道组具有更高的脂肪酸含量, 其中包括丰富的不饱和脂肪酸, 如C20:1花生烯酸、C16:1棕榈油酸、C18:3n6 γ-亚油酸、C18:2n6c亚油酸、C18:1n9c油酸和C18:3n3 α-亚油酸等。相反, 池塘组的粗脂肪成分可能主要由类脂(例如固醇和磷脂)组成。有研究指出更高的不饱和脂肪酸以及相对较低的固醇含量, 有助于缓解炎症和心血管疾病(Liu et al, 2023; Simonen et al, 2023), 从而使跑道组鲫鱼更适合健康饮食模式。此外GC-IMS分析获得的挥发性风味成分占33种, 占总指标的55%。这意味着在两种养殖模式下, 鲫鱼的挥发性成分存在显著差异。这些差异可能源自不同养殖模式下鱼类生长环境、饲料和代谢产物的变化, 进一步影响了其品质特征。因此, 相较于其他指标检测分析需要更多的时间成本, GC-IMS分析可用于快速区分不同养殖模式下鲫鱼的挥发性风味物质差异, 从而在食品鉴别方面具有巨大潜力。总之, 不同的养殖模式对鲫鱼的营养价值和风味产生巨大影响。本研究对水产品养殖模式选择、产品定位以及食品质量控制提供了重要的科学支持。
 |
| 图 4 不同养殖模式下鲫鱼的层次聚类热图 Fig. 4 Hierarchical clustering heatmaps of C. auratus from different aquaculture modes |
通过分析两种养殖模式鲫鱼的各项指标发现, 两组鲫鱼在不同养殖方式下的形体确有差异, 且跑道组鲫鱼呈现出高蛋白低脂肪的特点。两组鲫鱼的蒸煮损失和持水力均无显著差异, 但跑道组鲫鱼肌肉蛋白质更易于人体消化。此外, 跑道组鲫鱼的大部分游离氨基酸含量都高于池塘组, 且跑道组鲫鱼呈现鲜甜味的氨基酸含量更显著, 因此跑道组鲫鱼比池塘组鲫鱼可能具有更好地风味。两组鲫鱼肌肉氨基酸均符合FAO/WHO的理想蛋白质模式, 含有丰富氨基酸且组成合理均衡。同时跑道组鲫鱼的脂肪酸组成更合理均衡, 营养价值更高, 可作为人体的优质脂肪来源。根据GC-IMS分析和层次聚类分析的结果, 验证了不同养殖模式的鲫鱼中存在着挥发性风味物质的差异, 且跑道组鲫鱼中的1-辛烯-3-醇和三甲胺含量较低, 这表明跑道组鲫鱼可能具有更佳的风味质量, 同时也说明GC-IMS技术可快速有效鉴定不同养殖模式下的鲫鱼。综上表明, 池塘内循环流水跑道养殖在一定程度上可改善鲫鱼营养品质和特征风味, GC-IMS技术在鉴定不同养殖模式鱼类方面潜力巨大, 同时也为消费者日常选购、养殖者养殖模式选择以及产品定位等方面提供了理论依据。
王珂欣, 贾京华, 刘薇, 等, 2023. 超声辅助水酶法提取赤子爱胜蚓油脂及其脂肪酸组成分析[J/OL]. 中国油脂, 1-9.
(2023-06-16). https://doi.org/10.19902/j.cnki.zgyz.1003-7969.230147.
|
史庆超, 熊小琴, 王均, 等, 2021. 高位池和传统池塘养殖大口鲶肌肉营养品质的比较[J]. 现代畜牧兽医, (2): 35-38. |
朱士臣, 冯媛, 彭建, 等, 2021. 池塘内循环水"跑道"系统与传统池塘养殖的三角鲂营养品质对比研究[J]. 食品安全质量检测学报, 12(3): 1057-1065. |
刘凯, 李奎, 石伟, 等, 2021. 池塘内循环流水养殖草鱼的研究[J]. 湖南农业科学, (8): 70-73. |
刘梅, 宓国强, 郭建林, 等, 2019. 池塘内循环流水养殖模式对黄颡鱼生长性能、形体指标、血清生化指标及肌肉营养成分的影响[J]. 动物营养学报, 31(4): 1704-1717. |
刘梅, 原居林, 倪蒙, 等, 2021. 大口黑鲈(Micropterus salmoides)不同养殖模式氮磷收支及养殖效果研究[J]. 海洋与湖沼, 52(3): 718-728. |
孙立春, 陈丽娟, 倪玲, 等, 2012. 纸型海藻食品的氨基酸构成和模拟胃肠消化特性的研究[J]. 食品科学, 33(19): 141-145. |
孙慧娟, 李璐, 马凯华, 等, 2023. 革胡子鲶鱼皮营养成分和挥发性物质分析[J]. 中国食品学报, 23(2): 341-352. |
李双琦, 崔震昆, 2022. 热处理方式对鲈鱼游离氨基酸及品质影响[J]. 中国调味品, 47(1): 32-35, 45. |
李倩, 郭建林, 王雨辰, 等, 2020. 池塘内循环流水养殖下太湖鲂鲌(翘嘴鲌(♀)×三角鲂(♂))肠道微生物群落变化的研究[J]. 海洋与湖沼, 51(2): 364-370. |
李敏, 蔡景波, 张翔, 等, 2022. 2种养殖模式梭鱼肌肉营养组成及挥发性风味成分比较[J]. 浙江海洋大学学报(自然科学版), 41(2): 119-126, 148. |
吴俊杰, 魏帮鸿, 郭全友, 等, 2023. 即食烤制鱿鱼工艺优化及风味分析[J/OL]. 食品与发酵工业, 1-11. (2023-06-16).
https://doi.org/10.13995/j.cnki.11-1802/ts.035306.
|
赵亭亭, 张岩, 陈超, 等, 2018. 3种养殖石斑鱼的肌肉营养成分分析与品质评价[J]. 渔业科学进展, 39(6): 89-96. |
姜晓凡, 2021. 棕榈油酸通过SIRT3调控高脂诱导的肝脏糖异生的作用及其机制[D]. 济南: 山东大学.
|
贾丽, 潘勇, 刘帅, 2011. 池塘内循环流水养殖模式——美国的一种新型养殖模式[J]. 中国水产, (1): 40-42. |
徐永霞, 白旭婷, 冯媛, 等, 2021. 基于GC-IMS和化学计量学分析海鲈鱼肉蒸制过程中风味物质的变化[J]. 食品科学, 42(22): 270-275. |
高山, 王嘉佳, 胡高爽, 2020. 沙棘果油棕榈油酸提取物的富集及改善胰岛素抵抗活性[J]. 食品科学, 41(10): 272-280. DOI:10.7506/spkx1002-6630-20190429-381 |
黄爱霞, 陈建明, 姜建湖, 等, 2021. 池塘内循环跑道养殖青鱼形体指数和肌肉营养价值分析[J]. 科学养鱼, (5): 73-74. DOI:10.3969/j.issn.1004-843X.2021.05.039 |
董立学, 喻亚丽, 毛涛, 等, 2021. 池塘内循环流水养殖斑点叉尾 |
蔡振鑫, 刘春红, 2021. 基于神经网络的多特征结合法鲫鱼质量估计[J]. 中国农业科技导报, 23(12): 109-115. |
AASLYNG M D, MEINERT L, 2017. Meat flavour in pork and beef – From animal to meal[J]. Meat Science, 132: 112-117. DOI:10.1016/j.meatsci.2017.04.012 |
CAI W J, LIU H K, HE L Y, et al, 2023. Exercise training combined with a high-fat diet improves the flesh flavour, texture and nutrition of gibel carp (Carassius auratus gibelio)[J]. Food Chemistry: X, 17: 100612. DOI:10.1016/j.fochx.2023.100612 |
DING M Z, HUANG Z X, JIN Z Y, et al, 2022. The effect of fat content in food matrix on the structure, rheological properties and digestive properties of protein[J]. Food Hydrocolloids, 126: 107464. DOI:10.1016/j.foodhyd.2021.107464 |
DUAN Z L, DONG S L, DONG Y W, et al, 2021. Geographical origin identification of two salmonid species via flavor compound analysis using headspace-gas chromatographyion mobility spectrometry combined with electronic nose and tongue[J]. Food Research International, 145: 110385. DOI:10.1016/j.foodres.2021.110385 |
FRITSCHE K L, 2015. The science of fatty acids and inflammation[J]. Advances in Nutrition, 6(3): 293S-301S. DOI:10.3945/an.114.006940 |
HEISING J K, VAN BOEKEL M A J S, DEKKER M, 2014. Mathematical models for the trimethylamine (TMA) formation on packed cod fish fillets at different temperatures[J]. Food Research International, 56: 272-278. DOI:10.1016/j.foodres.2014.01.011 |
LIU Y, SHEN N, XIN H W, et al, 2023. Unsaturated fatty acids in natural edible resources, a systematic review of classification, resources, biosynthesis, biological activities and application[J]. Food Bioscience, 53: 102790. DOI:10.1016/j.fbio.2023.102790 |
MARTINS N, MAGALHÃES R, VIERIA L, et al, 2023. Dietary oleic acid supplementation improves feed efficiency and modulates fatty acid profile and cell signaling pathway in European sea bass (Dicentrarchus labrax) juveniles fed high-lipid diets[J]. Aquaculture, 576: 739870. DOI:10.1016/j.aquaculture.2023.739870 |
SIMONEN P, ÖÖRNI K, SINISALO J, et al, 2023. High cholesterol absorption: A risk factor of atherosclerotic cardiovascular diseases?[J]. Atherosclerosis, 376: 53-62. DOI:10.1016/j.atherosclerosis.2023.06.003 |
VASKOSKA R, HA M, ONG L, et al, 2021. Myosin sensitivity to thermal denaturation explains differences in water loss and shrinkage during cooking in muscles of distinct fibre types[J]. Meat Science, 179: 108521. DOI:10.1016/j.meatsci.2021.108521 |
WANG S Q, CHEN H T, SUN B G, 2020. Recent progress in food flavor analysis using gas chromatography–ion mobility spectrometry (GC-IMS)[J]. Food Chemistry, 315: 126158. DOI:10.1016/j.foodchem.2019.126158 |
ZHANG L H, XU N, LIU X L, et al, 2021. Exercise training accelerates UPS- and mTOR-mediated protein turnover of grass carp Ctenopharyngodon idella[J]. Aquaculture, 545: 737252. DOI:10.1016/j.aquaculture.2021.737252 |
ZHANG T T, XU J, WANG Y M, et al, 2019. Health benefits of dietary marine DHA/EPA-enriched glycerophospholipids[J]. Progress in Lipid Research, 75: 100997. DOI:10.1016/j.plipres.2019.100997 |
ZHAO D D, HU J, CHEN W X, 2022. Analysis of the relationship between microorganisms and flavour development in dry-cured grass carp by high-throughput sequencing, volatile flavour analysis and metabolomics[J]. Food Chemistry, 368: 130889. DOI:10.1016/j.foodchem.2021.130889 |