 2024, Vol. 55
2024, Vol. 55中国海洋湖沼学会主办。
文章信息
- 张帆, 郑关超, 王潇潇, 翟毓秀, 谭志军, 吴海燕. 2024.
- ZHANG Fan, ZHENG Guan-Chao, WANG Xiao-Xiao, ZHAI Yu-Xiu, TAN Zhi-Jun, WU Hai-Yan. 2024.
- 基于UPLC-Q-Exactive MS构建麻痹性贝类毒素的特征指纹溯源技术
- CONSTRUCTION OF A FINGERPRINT TRACEABILITY TECHNIQUE BASED ON UPLC-Q-EXACTIVE MS FOR THE CHARACTERIZATION OF PARALYTIC SHELLFISH POISON
- 海洋与湖沼, 55(1): 135-144
- Oceanologia et Limnologia Sinica, 55(1): 135-144.
- http://dx.doi.org/10.11693/hyhz20230500110
文章历史
-
收稿日期:2023-05-19
收修改稿日期:2023-08-05




2. 中国海洋大学食品科学与工程学院 山东青岛 266003;
3. 海水养殖生物育种与可持续产出全国重点实验室 中国水产科学研究院黄海水产研究所 山东青岛 266071
2. Collage of Food Sciences & Technology, Ocean University of Cina, Qingdao 266003, China;
3. State Key Laboratory of Mariculture Biobreeding and Sustainable Goods, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
麻痹性贝类毒素(paralytic shellfish toxins, PSTs)主要由Alexandrium、Gymnodinium和Pyrodinium等海洋甲藻产生, 通过贝类滤食进入食物链, 在世界范围内造成严重生态和食品安全风险, 是国际社会重点关注目标和管控对象(Visciano et al, 2016; Nicolas et al, 2017)。研究表明PSTs分布具有地域性特点(Villalobos et al, 2019), 而其风险形成过程主要受两个方面的影响: 首先是产毒藻的种类及来源差异, 链状亚历山大藻(Alexandrium catenella)主要分布在渤海、黄海和东海海域(顾海峰等, 2011; 唐莹莹等, 2018), 而太平洋亚历山大藻主要分布在东海和南海, 链状裸甲藻在福建近海曾多次导致中毒事件(于仁成等, 2020)。其次, 不同地区贝类如紫贻贝摄食产毒藻后毒素成分也有显著差异, 在2019年大连大窑湾海域紫贻贝中PSTs成分以C1 & 2和GTX2 & 3为主(许道艳等, 2014), 而在浙南海域的紫贻贝中PSTs成分则以C2、GTX5和NEO为主(张树刚等, 2011), 这种区域差异在其他研究中也得到了充分的证明(Yao et al, 2019; Liang et al, 2022)。因此, 贝类PSTs组分的区域差异对其风险具有一定的指示作用, 可有效指向风险源地并用于产地溯源, 对于提升贝类中PSTs风险的监管效果具有重要作用。
近年来, 指纹溯源技术广泛应用于生物地球化学、医学领域、环境监测、食品安全等多个领域, 通过对生物体指纹生物标志物的筛选, 结合数据库比对可快速实现源头识别(刘静等, 2022; 冷雪等, 2023)。该技术是基于代谢组学技术, 以高通量、高灵敏度、高分辨率的现代仪器分析方法为手段, 对细胞、体液和组织中全部代谢物进行定性与定量分析, 随后结合多元统计分析来构建指纹溯源技术模型(孙晓珊等, 2021; 高淑芳等, 2022)。其中液相色谱-质谱联用(LC-MS)(徐天润等, 2020)能够在任何单一分析平台中实现最高的代谢组覆盖率, 并且需要最少的样品前处理, 包括蛋白质沉淀和代谢物提取(Bujak et al, 2015), 适宜的提取溶剂、提取时间和处理温度等条件是影响前处理的重要参考指标。有文献报道, 基于LC-MS的代谢指纹识别鲤鱼和虹鳟鱼精浆中的代谢物, 筛选出精子质量的新型生物标志物, 为优化人工繁殖奠定了基础(Dietrich et al, 2019)。同时也有研究针对澳大利亚翡翠贻贝新西兰绿唇贻贝和进行代谢指纹检测, 发现两种贻贝之间存在明显的代谢差异, 根据差异代谢物可以实现贻贝产地溯源(Rochfort et al, 2013)。此外, 也有研究采用同位素标记(Zhao et al, 2019)、脂质(Shin et al, 2008)、微量元素(Morrison et al, 2019)等技术识别双壳贝类肠道内容物的方式, 实现贝类产地溯源(Gao et al, 2006)。
采用高通量筛查技术获得的数据规模大且来源复杂, 因此需要结合化学计量学方法, 降低数据维度筛选出具有识别能力的变量(徐天润, 2020)。多变量分析是通过降低维度来简化数据的复杂程度, 并通过相关软件进行结果可视化, 主要包括主成分分析(principal component analysis, PCA)、正交偏最小二乘判别分析(orthogonal signal correction partial least squares-discriminant analysis, OPLS-DA)、聚类分析等。其中PCA主要通过质量控制样本(QC)在PCA图中分布情况对数据进行初步考察, 谭芷晴等(2023)为筛选炔诺酮暴露斑马鱼后的生物标志物, 运用PCA分析发现处理组与代谢组有明显分离。刘天亮等(2022)运用PCA分析成功将来自山东、河南和河北产区的金银花样品分成了3类。而OPLS-DA可以使分类信息主要集中在主成分上, 并且通过变量投影重要度(Variable importance in projection, VIP)来确定差异化合物, 判别效果更具有说服力(李思源等, 2021)。然而, 基于多变量分析的代谢指纹从未用于研究摄食麻痹性贝类毒素的紫贻贝。
本研究选择我国近海主要麻痹性贝类毒素产毒藻种, 通过室内暴露贻贝模拟指纹信息传递过程, 采用UPLC-Q-Exactive MS对五种前处理方法进行比较分析, 结合PCA和OPLS-DA化学计量学方法, 挖掘摄食不同产毒藻后贻贝体内的化学差异, 筛选区分摄食不同产毒藻贻贝中的特征指纹物质, 构建特征指纹溯源技术模型, 快速精准的识别潜在风险以便提高麻痹性贝类毒素风险源头识别。
1 材料与方法 1.1 实验材料实验所用藻株包括链状亚历山大藻(A. catenella, GY-H25)、微小亚历山大藻(A. minutum, GY-H46)和链状裸甲藻(G. catenatum, GY-H65)购自上海光语生物科技有限公司。实验室内以f/2培养液单种培养, 温度为(20±1) ℃, 光照为6 000 lx; 光暗比12 h︰12 h。饵料藻选用小球藻(Chlorella vulgaris), 置于−20 ℃冰箱保存。
样品前处理及色谱质谱条件优化采用取自秦皇岛贻贝重点养殖区的紫贻贝(Mytilus galloprovincialis), 经检测为麻痹性贝类毒素阳性贝, 毒素含量为1 008 µg STXeq/kg。
指纹溯源技术建立采用购于山东青岛码头紫贻贝(Mytilus galloprovincialis), 平均规格为: 壳长(40.0±1.9) mm, 壳宽(22.6±0.8) mm, 总重(6.0±0.6) g, 购入后先在实验室进行驯化, 每天持续通气并更换海水。实验共设置3组, 每200只紫贻贝为一组, 培养于24 h通气的天然海水养殖容器环境下。
1.2 实验方法 1.2.1 紫贻贝室内暴露实验各取1 mL混匀后的链状亚历山大藻、微小亚历山大藻和链状裸甲藻的藻液, 加入20 μL鲁哥试剂(Lugol’s agent)固定, 在显微镜下计数藻细胞密度。按照A. catenella组和A. minutum组投喂量约为1×105 cells/(inds.·d), G. catenatum组投喂量约为2×105 cells/(inds.·d), 早晚各喂一次, 轻微曝气。实验共持续14 d, 投喂产毒藻7 d, 投喂饵料藻7d。分别取0 (对照)、1、3、7和14 d的紫贻贝样品, 每组样品设置3个生物学重复, 毒素分析样品为3只贝混样; 特征物质分析样品为6只贝混样, 均解剖紫贻贝并采集全部软组织, 于−80 ℃下冷冻保存。
1.2.2 产毒藻和贻贝中麻痹性贝类毒素组成分析将指数生长期的藻液取20 mL于离心管中, 2 760 ×g离心5 min, 收集藻细胞置于15 mL离心管中, 加入5 mL 1%乙酸水溶液。将其置于冰浴中, 使用超声波细胞破碎仪(JY92-ⅡN, 宁波新芝生物科技股份有限公司)对藻类样品进行破碎(设置全程时间5 min、间隔时间2 s、功率比50%)。镜检确认藻细胞完全破碎后, 将离心管置于高速冷冻离心机(Himac CR 22G, 日本Hitachi公司)中, 2 760 ×g离心5 min, 供测试。将贻贝开壳用生理盐水清洗后取软组织, 沥水均质, 参考Boundy等(2015)等方法进行麻痹性贝类毒素提取及测试。
1.2.3 特征指纹物质的前处理方法取(1.00±0.02) g麻痹性贝类毒素阳性紫贻贝样品转移至5 mL离心管(分为5组, 每组6个平行), 按照表 1中五种方法进行前处理, 提取液经12 470 ×g离心5 min后, 供测试。
| 方法 | 提取剂 | 加速提取方法 | 净化方法 |
| 方法A (仲云, 2020) |
甲醇/水(体积比为1︰1, 含0.1%乙酸) | 超声30 min | 无 |
| 方法B (Wang et al, 2022) |
甲醇/乙腈/水(体积比为2︰2︰1) | 液氮中冷冻1 min, 冰水中4 ℃超声10 min, 重复上述步骤三遍 | 提取液−20 ℃冰箱放置1 h去除蛋白, 离心浓缩后100 µL乙腈/水(体积比为1︰1)复溶 |
| 方法C (Gallon et al, 2021) |
甲醇/水(体积比为7︰3) | 冰水浴超声10 min | 无 |
| 方法D (Dong et al, 2020) |
乙腈/甲醇/丙酮(体积比为1︰1︰1) | 涡旋15 min | 30 ℃氮气干燥, 在1 mL甲醇中重新 溶解 |
| 方法E (Letsinger et al, 2020) |
甲醇/氯仿(体积比为2︰1) | 涡旋20 min, 4 416 ×g离心, 二次提取, 合并上清液, 加入600 µL冰水, 3 312 ×g离心, 得到相分离。 | 上相通过0.22 µm的过滤器, 然后氮吹。浓缩样品用1 mL甲醇/水(体积比为1︰1)重新溶解 |
Q-Exactive液相色谱质谱联用仪(Q-Exactive, 美国Thermo公司)C8色谱柱体系: Kinetex C8 (150 mm×2.1 mm, 2.6 µm); 梯度洗脱程序: 0~0.5 min, 20% A; 0.5~5.0 min, 20%~90% A; 5.0~14.0 min, 90% A; 14.0~15.0 min, 80% A。Amide色谱柱体系: TSK-Amide-80柱(2 mm×15 cm, 3 μm); 梯度洗脱程序: 0~1.0 min, 80% A; 1.0~4.0 min, 80%~ 40% A; 4.0~6.5 min, 40% A; 6.5~8.0 min, 40%~80%A; 8.1~10.0 min, 80% A。
柱温: 40 ℃; 进样量: 10 μL; 流速: 0.35 mL/min; 流动相为: A相为95%乙腈水(含2 mmol/L甲酸铵, 50 mmol/L甲酸), B相为水(含2 mmol/L甲酸铵, 50 mmol/L甲酸)。
1.2.5 质谱条件Q-Exactive液相色谱质谱联用仪使用加热电喷雾离子源(HESI), 扫描模式: 全扫描/数据依赖二级子离子扫描(Full MS/dd MS2); 喷雾电压: 3 500 V; 毛细管和加热器温度: 320和50 ℃; 鞘气和辅助气: 40和10 arb; 分辨率用半峰高处的全峰宽(FWHM)表示, 一级全扫描分辨率: 70 000 FWHM; trap最大容量(AGC target): 1×106; C-trap最大注入时间: 150 ms, 数据依赖二级离子扫描分辨率: 17 500 FWHM; C-trap最大容量(AGC target): 1×105; C-trap最大注入时间: 50 ms; 质谱扫描范围: 质荷比(m/z) 100~600和600~1 500。
1.3 数据处理 1.3.1 样品前处理及色谱质谱分析按照时间顺序在色谱图中随机选取峰强度大于1×105的10个离子峰, 分别计算每种方法6个样品的峰强度均值, 计算每种方法贻贝样品中所测的10个离子峰峰强度及其变异系数(Coefficient of variation, CV=标准差/均值)。5种前处理方法所测峰强度以均值表示, 多组间比较采用随机区组的单因素方差分析, 若组间差异有统计学意义, 进一步的两两比较采用Bonferroni检验。以P < 0.05为差异有统计学意义。
1.3.2 特征指纹物质分析原始数据使用Compound Discoverer 3.1软件进行峰检测、保留时间对齐等预处理。采用数据库(ChemSpider, mzCloud等)从精确质量数、同位素组成及分布与预测分子式的吻合度、一级质谱/二级质谱碎片和数据库匹配等方面进行化合物初步鉴定, 其中设置参数如下: 保留时间最大漂移值0.2 min; 质量偏差<5×10−6; 信号强度偏差<30%; 峰强度最小值度50 000, 同时设置基质空白去除, 不进行统计学分析。由于特征物质没有标准品, 采用STX标准品曲线进行相对定量分析计算其浓度。
1.4 统计分析采用Excel 2021、Origin 2023、SIMCA 14.1和IBM SPSS Statistics 22软件对数据进行统计分析和绘图。在Excel 2021中将数据整理为二维矩阵形式, 保留P值、m/z值、保留时间、分子式、峰面积等信息, 根据P值筛选出差异显著的化合物(P < 0.01); 以化合物的峰面积为变量, m/z值作为观测ID将数据导入SIMCA 14.1进行PCA与OPLS-DA分析; 根据VIP > 1作为特征指纹物质。采用逐步判别法对3组样品的13个复合指纹物质特征建立3个群体的Fisher线性判别函数。
2 结果与分析 2.1 样品前处理及色谱质谱条件选择通过对比不同色谱和质谱条件结果如图 1所示, 两种色谱柱均为方法D检测到的化合物数量最多。采用C8色谱柱时(图 1a), 正离子模式m/z 100~600扫描范围下检测到的化合物数量最多为3 374个, 负离子模式m/z 100~600扫描范围次之; 采用Amide色谱柱时(图 1b)负离子模式m/z 100~600的扫描范围下检测到的化合物最多为1 785个, 正离子模式m/z 600~ 1 500扫描范围次之。因此方法D检测到的化合物达到了6 811个, 更加适合对特征物质的提取。
 |
| 图 1 不同前处理方法在每种色谱质谱条件下的特征物质数量 Fig. 1 The amount of compounds in each mode for different pretreatment method a: C8色谱柱; b: Amide色谱柱 |
此外, 根据不同方法的峰强度变异系数(CV值)来分析不同的质谱条件。采用C8色谱柱时, 离子峰强均值在正离子模式m/z 100~600和m/z 600~1 500扫描范围下大于1×107且差异无统计学意义(P > 0.05); 比较其CV值, 方法D在正离子模式m/z 600~1 500扫描范围下CV值最小。采用Amide色谱柱时, 方法D在正离子模式m/z 600~1 500扫描范围下的峰强最高为5.49×107且差异具有统计学意义(P < 0.05), CV值< 25% (表 2)。因此, 选择方法D在C8色谱柱正离子模式m/z 100~600的扫描范围和Amide色谱柱正离子模式m/z 600~1 500的扫描范围下更适用于贻贝的特征物质分析, 覆盖的化合物数量占总检测量的40.4%。在方法D两种色谱质谱条件下, 分别在QC样品色谱图的前、中、后随机共选取10个离子峰进行分析, 其CV值均≤1.00%, 各峰面积和峰强度的CV值均<25.00%。以上结果表明本研究样品分析过程中仪器的精密度、稳定性及重复性均较好, 满足分析要求。
| 色谱柱体系 | 扫描范围m/z | 扫描模式 | 峰强 | CV/% | |||||||||
| 方法A | 方法B | 方法C | 方法D | 方法E | 方法A | 方法B | 方法C | 方法D | 方法E | ||||
| Amide | 100~600 | 正离子 | 9.40×107 | 8.72×107 | 1.56×106 | 2.11×106 | 2.22×106 | 26.3 | 49.5 | 59.6 | 61.6 | 64.7 | |
| 负离子 | 1.63×107 | 1.72×107 | 1.95×107 | 1.39×107 | 1.50×107 | 32.2 | 39.5 | 41.1 | 30.4 | 81.0 | |||
| 600~1 500 | 正离子 | 4.98×107 | 5.42×107 | 4.61×107 | 5.49×107 | 4.53×107 | 8.97 | 23.3 | 13.1 | 16.0 | 54.7 | ||
| 负离子 | 9.44×106 | 9.30×106 | 5.23×106 | 7.47×106 | 1.05×107 | 22.0 | 10.1 | 16.8 | 12.3 | 12.0 | |||
| C8 | 100~600 | 正离子 | 3.88×108 | 4.64×108 | 2.94×108 | 8.02×108 | 6.20×108 | 30.9 | 72.4 | 22.4 | 9.91 | 17.6 | |
| 负离子 | 2.47×108 | 4.06×108 | 2.77×108 | 8.01×108 | 5.80×108 | 33.9 | 77.6 | 23.7 | 11.0 | 19.5 | |||
| 600~1 500 | 正离子 | 6.11×106 | 1.42×106 | 6.43×107 | 2.50×107 | 2.55×107 | 54.4 | 105 | 19.4 | 32.6 | 34.5 | ||
| 负离子 | 6.53×106 | 1.68×106 | 1.99×107 | 1.34×107 | 4.07×107 | 73.8 | 51.6 | 64.8 | 28.2 | 44.6 | |||
紫贻贝摄食不同产毒藻后麻痹性贝类毒素的变化情况如图 2所示, 在前7天为毒素蓄积阶段, 紫贻贝中的麻痹性贝类毒素含量不断上升, 由暴露1天毒素含量最低为54.1 µg STXeq/kg, 到7天时毒素含量最高为5 538 µg STXeq/kg; 7天到14天为代谢阶段(停止投喂产毒藻), 至14天时毒素含量最低降至267 µg STXeq/kg。
 |
| 图 2 紫贻贝摄食不同产毒藻后体内麻痹性贝类毒素含量及组成 Fig. 2 Variation of toxin content and composition after ingestion of different toxin producing algae by M. galloprovincialis 注: a: 链状亚历山大藻(A. catenella); b: 微小亚历山大藻(A. minutum); c: 链状裸甲藻(G. catenatum) |
在投喂产毒藻的紫贻贝体内共检出11种麻痹性贝类毒素成分, 不同组样品毒素成分差异显著。投喂A. catenella的紫贻贝中麻痹性贝类毒素主要成分为C1 & 2和GTX5, 在整个实验中C1 & 2占比从第1天68.6%下降到第14天的49.8%; 投喂A. minutum的紫贻贝中麻痹性贝类毒素主要成分为GTX1 & 4和GTX2, 且在整个实验期间各成分占比稳定; 投喂G. catenatum的紫贻贝中麻痹性贝类毒素主要成分为dcGTX2 & 3、dcSTX和C1 & 2, 其中dcSTX占比从第1天的10.9%增高到第14天的21.5%; 而C2的占比从21.5%降至4.02%。因此, 摄食不同产毒藻后紫贻贝体内的毒素成分有明显差异且随时间发生代谢转化, 还需对紫贻贝体内的代谢物进行检测。
2.3 多元统计分析 2.3.1 主成分分析本研究使用上述乙腈/甲醇/丙酮(体积比为1︰1︰1)作为提取剂的提取方法结合筛选出的两个离子通道, 经过软件处理后, 提取了3 886个化合物, 利用P < 0.01筛选差异化合物并结合毒素成分进行PCA分析, 如图 3所示, 前两个主成分分别解释了总变量的36.9%和18.3%, 共解释了55.2%的变量。三组样品分别沿PC1或PC2轴分离, 表明摄食不同产毒藻贻贝中化合物存在差异。
 |
| 图 3 投喂三种不同产毒藻后贻贝中差异化合物PCA图 Fig. 3 PCA of different compounds and toxins in M. galloprovincialis fed with three different toxic microalgae |
对投喂A. catenella、A. minutum和G. catenatum的贻贝经筛选后结合毒素进行OPLS-DA分析, 如图 4a~4c所示。三组样品中的化合物显著分离, 自变量拟合指数(RX2)分别为0.724、0.860、0.846, 自变量拟合指数(RY2)分别为1.00、1.000、0.969, 模型预测指数(Q2)为0.985、0.982、0.988, 表明当前OPLS-DA模型稳定可靠, 具有良好的预测性。经OPLS-DA模型分析分别生成S-plots图, 如图 4d~4f所示。OPLS-DA模型中变量VIP > 1.0说明该变量对整体模型的贡献度高于平均水平, 作为本研究的毒素和特征指纹物质。此外, 毒素成分NEO和dcNEO作为摄食A. catenella和G. catenatum后紫贻贝的特有成分也被作为指纹物质, 共计13种复合指纹物质。根据精确分子质量、保留时间以及二级质谱碎片等与数据库进行比对参数如表 3所示。
 |
| 图 4 投喂三种不同产毒藻后贻贝中差异化合物OPLS-DA图和S-plots图 Fig. 4 OPLS-DA and S-plots of differential compounds and toxins in M. galloprovincialis fed with three different toxic microalgae 注: a~c: OPLS-DA图; d~f: S-plots图 |
| 类别 | 化学名称 | 分子质量/(g/mol) | 保留时间/min | 相对浓度/(ng/mL) | ||
| A. catenella | A. minutum | G. catenatum | ||||
| 特征指纹物质 | C17H31NO5 | 329.22 | 3.06 | 193 | 453 | 825 |
| C25H19NO3 | 381.14 | 4.66 | 5.64 | 19.0 | 26.5 | |
| C27H43NO5 | 461.31 | 5.02 | 479 | 396 | 1175 | |
| C24H26N2O2 | 374.21 | 4.74 | 514 | 578 | 1026 | |
| 毒素成分 | dcGTX2 | 353.09 | 5.50 | 5.04 | 18.8 | 129 |
| dcGTX3 | 353.09 | 5.66 | 1.91 | — | 76.0 | |
| GTX1 | 412.09 | 5.47 | 16.1 | 932 | — | |
| GTX4 | 412.09 | 5.57 | 6.76 | 264 | — | |
| GTX3 | 396.09 | 5.54 | 6.18 | 128 | — | |
| C2 | 476.05 | 5.48 | 225 | 0.00 | 57.1 | |
| STX | 300.14 | 6.03 | 15.8 | 17.8 | 14.3 | |
| NEO | 316.14 | 5.60 | 36.6 | — | — | |
| dcNEO | 273.13 | 5.07 | — | — | 38.5 | |
| 注: 特征指纹物质以STX曲线折算相对浓度; —表示未检出 | ||||||
根据筛选的复合指纹物质, 建立基于Fisher判别函数的一般判别方法。以筛选出来的13种复合指纹物质作为判别分析的自变量, 对贻贝样品进行多变量判别分析, 排除了7种对分类作用较小的指纹物质, 仅保留6种指纹物质用于判别分析, 建立Fisher线性判别函数如下:
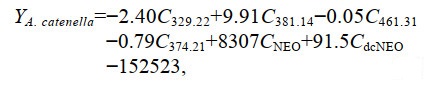 (1)
(1)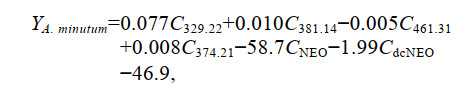 (2)
(2)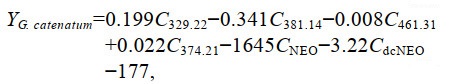 (3)
(3)式中: Y分别表示暴露于不同产毒藻贻贝的判别得分; C329.22、C381.14、C461.31、C374.21、CNEO、CdcNEO分别表示分子质量为329.22、381.14、461.31、374.21的特征指纹物质和毒素NEO、dcNEO。回代检验正确判别率为100%, 交叉验证正确率为88.9%, 表明6种特征指纹成分对贻贝的判别效果较好。
3 讨论麻痹性贝类毒素的风险形成过程, 亦是从产毒藻信息向贝类传递的过程。通过传统的监管手段不仅耗费大量的人力物力财力, 对于近年来常见的小规模有害赤潮爆发作用有限(Fernandes-Salvador et al, 2021)。研究利用UPLC-Q-Exactive MS结合PCA分析与OPLS-DA分析, 从摄食3种麻痹性贝类毒素产毒藻的紫贻贝样品里共筛选出13种复合指纹物质, 对摄食不同藻的紫贻贝进行成功溯源。由于有毒藻种类和生态学特征不同, 其所产PSTs的种类和含量也有很大差异(Gu et al, 2013), 因此造成了贝类中PSTs风险表征差异明显, 这为识别PSTs风险源头提供了重要依据。Lewis等(2022)用苏格兰的链状亚历山大藻和英格兰南部的链状亚历山大藻分别投喂紫贻贝测定其体内的毒素谱, 根据毒素谱差异成功区分了摄食两种产毒藻的样品。同时有研究(Yan et al, 2022)表明不同产毒藻的毒素组成及毒性大小存在着较大差别, 并且我国的毒素分布也具有区域性特征。由于双壳贝类摄食产毒藻后毒素会在体内发生代谢转化, 如摄食链状亚历山大藻后的紫贻贝中毒素成分随时间发生转化, C2占比减小而GTX5占比增加(张海涛等, 2023), 蛤仔在摄食微小亚历山大藻后毒素的积累阶段, GTX1 & 4的占比逐渐增加, 在排出阶段占比下降(焦玥等, 2010), 这与本研究结果一致。在之前研究中发现这种代谢转化会受到投喂产毒藻量(Chen et al, 2001)、环境的酸碱性(Che et al, 2020)等因素影响, 因此仅根据毒素信息无法进行准确分类。
研究表明, 含水的有机提取剂如甲醇水溶液(Sidwick et al, 2017)可以有效地提取极性代谢物, 而非极性代谢物常选择氯仿、二氯甲烷等有机试剂(Cao et al, 2020)进行提取。通过比较了5种高效前处理方法, 经过扣除空白贻贝基质干扰后, 最终选择了乙腈/甲醇/丙酮作为提取剂的提取方法, 可以有效地移除大分子蛋白质(Li et al, 2017), 提高弱极性和非极性物质(Mushtaq et al, 2014)的提取效率。通过比较, 特征物质的提取方法和色谱质谱方法差异系数较小, 表明仪器稳定较好(李倩倩等, 2021), 可为后续特异性物质的筛选提供了方法。采用PCA分析发现, 三组样品有明显分离; 进一步采用OPLS-DA分析, 剔除无关数据使得模型更加易于解释, 可视化效果更加明显(钟若梅等, 2018)。刘鸽等(2021)采用PCA与OPLS-DA分析, 通过VIP值> 1以及P < 0.05筛选出暴露无机砷后三疣梭子蟹的100种差异代谢产物。Vera等(2019)等采用主成分分析(PCA)并比较独立建模(SIMCA)和偏最小二乘判别分析(PLS-DA)作为化学计量学分类方法, 实现了对品种特级初榨橄榄油的地理来源高灵敏和无偏差的鉴定。进一步采用Fisher线性判别模型构建判别函数, 可在变量服从正态分布、没有显著相关或变量的平均值和方差不相关使效果较好(李洪成等, 2017)。欧阳建等(2022)通过对黄金茶绿茶风味的测定将52个样品分为醇厚型、鲜爽型和其他型3种滋味类型。开建荣等(2022)构建枸杞原产地鉴别的判别模型, 模型的正确判别率达到了97.3%。目前, 未见Fisher线性判别模型用于对摄食麻痹性贝类毒素产毒藻的溯源报道。本研究引入6种物质构建的判别模型交叉验证准确率达到了88.9%, 预测效果较为理想。指纹识别技术需要结合识别和鉴定(Cuadros-Rodríguez et al, 2021)两个过程, 为了达到更好地溯源目的还需要进一步的现场验证。同时, 由于在同一海域往往会存在多种产毒藻且毒素成分会更加复杂, 如辽东湾曾检测出安氏亚历山大藻、伪亚历山大藻和李氏亚历山大藻等(宋伦等, 2018), 长江口海域则为塔玛亚历山大藻和链状亚历山大藻(高岩等, 2016)。因此, 需持续采集典型藻株特征物质识别以增加溯源技术的准确度和适用性。
4 结论通过优化前处理方法, 确定了甲醇/乙腈/丙酮(体积比为1︰1︰1)作为提取剂; 采用Q-Exactive Orbitrap筛选C8色谱柱中正离子扫描模式m/z 100~600的扫描范围和Amide色谱柱正离子扫描模式m/z 600~1 500的扫描范围, 可获得40.4%的指纹信息。通过多元统计分析技术筛选出摄食不同产毒藻贻贝的13种复合指纹物质, 构建了判别模型交叉验证准确率达到了88.9%。研究成果有助于提高麻痹性贝类毒素区域风险源头的准确识别能力, 为防范食用安全风险提供了科学依据和技术支撑。
于仁成, 吕颂辉, 齐雨藻, 等, 2020. 中国近海有害藻华研究现状与展望[J]. 海洋与湖沼, 51(4): 768-788. |
开建荣, 石欣, 李彩虹, 等, 2022. 基于矿物元素技术的中宁不同产区枸杞的判别分析[J]. 食品与发酵工业, 48(9): 253-260. |
仲云, 2020. 一株利玛原甲藻的腹泻性贝类毒素产毒生理及制备技术研究[D]. 上海: 上海海洋大学.
|
刘天亮, 杨林林, 董诚明, 等, 2022. 基于化学模式识别的不同产地金银花HPLC指纹图谱研究[J]. 中草药, 53(15): 4833-4843. |
刘鸽, 李焕军, 张秀珍, 等, 2021. 基于代谢组学技术的无机砷对三疣梭子蟹鳃组织的毒性作用机制研究[J]. 中国水产科学, 28(5): 602-613. |
刘静, 亓超凡, 绪扩, 等, 2022. 指纹图谱技术在食品质量与安全中的应用研究进展[J]. 食品安全质量检测学报, 13(10): 3189-3197. DOI:10.3969/j.issn.2095-0381.2022.10.spaqzljcjs202210017 |
许道艳, 刘磊, 于姬, 等, 2014. 紫贻贝体内麻痹性贝毒在自然条件下的排出[J]. 海洋环境科学, 33(5): 705-708. |
孙晓珊, 路鑫, 许国旺, 2021. 基于高分辨质谱的代谢组学分析技术研究进展[J]. 质谱学报, 42(5): 787-803. |
李思源, 李培瑜, 刘弈彤, 等, 2021. 代谢组学在食品科学中的应用进展[J]. 食品与发酵工业, 47(5): 252-258. |
李洪成, 张茂军, 马广斌, 2017. SPSS数据分析实用教程[M]. 2版. 北京: 人民邮电出版社.
|
李倩倩, 任冠桦, 叶春华, 等, 2021. 基于液质联用的人血清代谢组学样品前处理方法研究[J]. 中国癌症防治杂志, 13(1): 75-80. DOI:10.3969/j.issn.1674-5671.2021.01.14 |
冷雪, 马艺鑫, 2023. 基于非靶向代谢组学的高脂血症大鼠肝脏和血清脂代谢异常相关生物标志物发现及人参皂苷Rb1干预机制研究[J]. 中国中药杂志, 48(14): 3922-3933. |
宋伦, 吴景, 李楠, 等, 2018. 辽东湾毒害微藻潜在风险分布及环境关联[J]. 中国环境科学, 38(8): 3049-3059. DOI:10.3969/j.issn.1000-6923.2018.08.034 |
张树刚, 刘亚林, 陈雷, 等, 2011. 浙南海域麻痹性贝毒研究[J]. 海洋环境科学, 30(5): 703-706. |
张海涛, 吴海燕, 郑关超, 等, 2023. 链状亚历山大藻暴露下紫贻贝体内麻痹性贝毒蓄积转化规律[J]. 渔业科学进展, 44(1): 181-190. |
欧阳建, 黄纯勇, 李适, 等, 2022. 黄金茶绿茶风味轮及滋味分类模型的构建[J]. 食品与发酵工业, 48(24): 139-146. |
钟若梅, 陈碧莹, 吴霞, 等, 2018. 基于主成分分析和正交偏最小二乘判别分析对NSAIDs干预RAW264.7细胞炎症模型的磷脂组学研究[J]. 广东药科大学学报, 34(1): 101-105. |
顾海峰, 刘婷婷, 蓝东兆, 2011. 中国沿海甲藻包囊研究进展[J]. 生物多样性, 19(6): 779-786. |
徐天润, 刘心昱, 许国旺, 2020. 基于液相色谱-质谱联用技术的代谢组学分析方法研究进展[J]. 分析测试学报, 39(1): 10-18. |
高岩, 于仁成, 柳阳, 等, 2016. 基于产毒基因sxtA的qPCR方法在长江口邻近海域有毒藻类检测中的应用初探[J]. 海洋环境科学, 35(2): 279-287. |
高淑芳, 张金鹏, 施永海, 等, 2022. 基于LC-MS技术的海、淡水养殖刀鲚卵巢的代谢组学比较分析[J]. 南方水产科学, 18(3): 68-75. |
唐莹莹, 乔玉宝, 蒋志伟, 等, 2018. 东海产麻痹性贝毒链状亚历山大藻共附生菌群多样性研究[J]. 海洋渔业, 40(6): 720-727. |
焦玥, 田华, 王宗灵, 等, 2010. 微小亚历山大藻投喂量与菲律宾蛤仔对麻痹性贝毒累积、转化和排出的动态关系[J]. 海洋科学进展, 28(2): 216-223. |
谭芷晴, 桂林, 李新昌, 等, 2023. 炔诺酮暴露下斑马鱼候选生物标志物筛选研究[J]. 生态科学, 42(2): 35-42. |
BOUNDY M J, SELWOOD A I, HARWOOD D T, et al, 2015. Development of a sensitive and selective liquid chromatography-mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction[J]. Journal of Chromatography A, 1387: 1-12. DOI:10.1016/j.chroma.2015.01.086 |
BUJAK R, STRUCK-LEWICKA W, MARKUSZEWSKI M J, et al, 2015. Metabolomics for laboratory diagnostics[J]. Journal of Pharmaceutical and Biomedical Analysis, 113: 108-120. DOI:10.1016/j.jpba.2014.12.017 |
CAO M S, HAN Q A, ZHANG J H, et al, 2020. An untargeted and pseudotargeted metabolomic combination approach to identify differential markers to distinguish live from dead pork meat by liquid chromatography-mass spectrometry[J]. Journal of Chromatography A, 1610: 460553. DOI:10.1016/j.chroma.2019.460553 |
CHE Y J, DING L, QIU J B, et al, 2020. Conversion and stability of new metabolites of paralytic shellfish toxins under different temperature and pH conditions[J]. Journal of Agricultural and Food Chemistry, 68(5): 1427-1435. DOI:10.1021/acs.jafc.9b07063 |
CHEN C Y, CHOU H N, 2001. Accumulation and depuration of paralytic shellfish poisoning toxins by purple clam Hiatula rostrata Lighttoot[J]. Toxicon, 39(7): 1029-1034. DOI:10.1016/S0041-0101(00)00242-7 |
CUADROS-RODRÍGUEZ L, ORTEGA-GAVILÁN F, MARTÍN-TORRES S, et al, 2021. Chromatographic fingerprinting and food identity/quality: potentials and challenges[J]. Journal of Agricultural and Food Chemistry, 69(48): 14428-14434. DOI:10.1021/acs.jafc.1c05584 |
DIETRICH M A, SAMCZUK P, CIBOROWSKI M, et al, 2019. Metabolic fingerprinting of carp and rainbow trout seminal plasma[J]. Aquaculture, 501: 178-190. DOI:10.1016/j.aquaculture.2018.11.017 |
DONG Y, QIU P, ZHAO L S, et al, 2020. Metabolomics study of the hepatoprotective effect of Phellinus igniarius in chronic ethanol-induced liver injury mice using UPLC-Q/TOF-MS combined with ingenuity pathway analysis[J]. Phytomedicine, 74: 152697. DOI:10.1016/j.phymed.2018.09.232 |
FERNANDES-SALVADOR J A, DAVIDSON K, SOURISSEAU M, et al, 2021. Current status of forecasting toxic harmful algae for the north-east Atlantic shellfish aquaculture industry[J]. Frontiers in Marine Science, 8: 666583. DOI:10.3389/fmars.2021.666583 |
GALLON M E, GOBBO-NETO L, 2021. Plant metabolites involved in the differential development of a Heliantheae-specialist insect[J]. Metabolites, 11(3): 134. DOI:10.3390/metabo11030134 |
GAO Q F, SHIN P K S, LIN G H, et al, 2006. Stable isotope and fatty acid evidence for uptake of organic waste by green-lipped mussels Perna viridis in a polyculture fish farm system[J]. Marine Ecology Progress Series, 317: 273-283. DOI:10.3354/meps317273 |
GU H F, ZENG N, LIU T T, et al, 2013. Morphology, toxicity, and phylogeny of Alexandrium (Dinophyceae) species along the coast of China[J]. Harmful Algae, 27: 68-81. DOI:10.1016/j.hal.2013.05.008 |
LETSINGER A C, MENON R, IYER A R, et al, 2020. A high fat/high sugar diet alters the gastrointestinal metabolome in a sex dependent manner[J]. Metabolites, 10(10): 421. DOI:10.3390/metabo10100421 |
LEWIS A M, DEAN K J, HARTNELL D M, et al, 2022. The value of toxin profiles in the chemotaxonomic analysis of paralytic shellfish toxins in determining the relationship between British Alexandrium spp. and experimentally contaminated Mytilus sp.[J]. Harmful Algae, 111: 102131. DOI:10.1016/j.hal.2021.102131 |
LI H, FAN S F, WANG Y, et al, 2017. Rapid detection of small molecule metabolites in serum of hepatocellular carcinoma patients using ultrafast liquid chromatography-ion trap-time of flight tandem mass spectrometry[J]. Analytical Sciences, 33(5): 573-578. DOI:10.2116/analsci.33.573 |
LIANG Y B, LI A F, CHEN J H, et al, 2022. Progress on the investigation and monitoring of marine phycotoxins in China[J]. Harmful Algae, 111: 102152. DOI:10.1016/j.hal.2021.102152 |
MORRISON L, BENNION M, GILL S, et al, 2019. Spatio-temporal trace element fingerprinting of king scallops (Pecten maximus) reveals harvesting period and location[J]. Science of the Total Environment, 697: 134121. DOI:10.1016/j.scitotenv.2019.134121 |
MUSHTAQ M Y, CHOI Y H, VERPOORTE R, et al, 2014. Extraction for metabolomics: access to the metabolome[J]. Phytochemical Analysis, 25(4): 291-306. DOI:10.1002/pca.2505 |
NICOLAS J, HOOGENBOOM R L A P, HENDRIKSEN P J M, et al, 2017. Marine biotoxins and associated outbreaks following seafood consumption: prevention and surveillance in the 21st century[J]. Global Food Security, 15: 11-21. DOI:10.1016/j.gfs.2017.03.002 |
ROCHFORT S J, EZERNIEKS V, MAHER A D, et al, 2013. Mussel metabolomics-Species discrimination and provenance determination[J]. Food Research International, 54(1): 1302-1312. DOI:10.1016/j.foodres.2013.03.004 |
SHIN P K S, YIP K M, XU W Z, et al, 2008. Fatty acid as markers to demonstrating trophic relationships among diatoms, rotifers and green-lipped mussels[J]. Journal of Experimental Marine Biology and Ecology, 357(1): 75-84. DOI:10.1016/j.jembe.2008.01.002 |
SIDWICK K L, JOHNSON A E, ADAM C D, et al, 2017. Use of liquid chromatography quadrupole time-of-flight mass spectrometry and metabonomic profiling to differentiate between normally slaughtered and dead on arrival poultry meat[J]. Analytical Chemistry, 89(22): 12131-12136. |
VERA D N, JIMÉNEZ-CARVELO A M, CUADROS-RODRÍGUEZ L, et al, 2019. Authentication of the geographical origin of extra-virgin olive oil of the Arbequina cultivar by chromatographic fingerprinting and chemometrics[J]. Talanta, 203: 194-202. |
VILLALOBOS L G, SANTINELLI N H, SASTRE A V, et al, 2019. Spatiotemporal distribution of paralytic shellfish poisoning (PSP) toxins in shellfish from Argentine Patagonian coast[J]. Heliyon, 5(6): e01979. |
VISCIANO P, SCHIRONE M, BERTI M, et al, 2016. Marine biotoxins: occurrence, toxicity, regulatory limits and reference methods[J]. Frontiers in Microbiology, 7: 1051. |
WANG R H, YIN Y D, LI J S, et al, 2022. Global stable-isotope tracing metabolomics reveals system-wide metabolic alternations in aging Drosophila[J]. Nature Communications, 13(1): 3518. |
YAN T, LI X D, TAN Z J, et al, 2022. Toxic effects, mechanisms, and ecological impacts of harmful algal blooms in China[J]. Harmful Algae, 111: 102148. |
YAO J Y, JIN W, LI D M, et al, 2019. Geographical distribution and seasonal variation in paralytic shellfish toxins in the coastal water of the South China Sea[J]. Toxicon, 168: 67-75. |
ZHAO X D, LIU Y, WANG G G, et al, 2019. Tracing the geographical origins of Yesso scallop (Patinopecten yessoensis) by using compound-specific isotope analysis: an approach for overcoming the seasonal effect[J]. Food Control, 102: 38-45. |