 2022, Vol. 53
2022, Vol. 53中国海洋湖沼学会主办。
文章信息
- 张孝民, 王秀霞, 李少文, 杨艳艳, 徐炳庆, 李凡. 2022.
- ZHANG Xiao-Min, WANG Xiu-Xia, LI Shao-Wen, YANG Yan-Yan, XU Bing-Qing, LI Fan. 2022.
- 基于优化模型的莱州湾三疣梭子蟹(Portunus trituberculatus)栖息地适宜性研究
- OPTIMIZATION OF HABITAT SUITABILITY MODEL FOR PORTUNUS TRITUBERCULATUS IN LAIZHOU BAY
- 海洋与湖沼, 53(3): 726-734
- Oceanologia et Limnologia Sinica, 53(3): 726-734.
- http://dx.doi.org/10.11693/hyhz20211100271
文章历史
-
收稿日期:2021-11-08
收修改稿日期:2021-12-22




栖息地适宜性是对栖息地特征与物种在该条件下生存质量的定量描述(易雨君等, 2013), 是当前海洋生态保护领域重要的研究内容之一, 对于生态养护、资源评估、种群动态研究都具有重要的理论和现实意义(邹易阳等, 2016)。在人类活动不断加剧和气候变化的大背景下, 全球生物多样性正处于持续的快速丧失中(徐炜等, 2016), 其中栖息地的减少已经成为物种和局部种群绝灭的重要原因(邓文洪, 2009; Keinath et al, 2017)。三疣梭子蟹(Portunus trituberculatus)生命周期短、繁殖力强、经济价值高(林群等, 2015), 捕捞数量占全世界经济蟹类的四分之一左右(Liu et al, 2013)。莱州湾位于渤海南部, 是赫赫有名的“莱州梭子蟹”的产地, 更有黄河和小清河等众多河流入海, 湾内基础饵料丰富, 水温盐度适宜, 是包括蟹类在内的众多经济性海洋生物的产卵和栖息场所。近几十年来, 受环境污染、过度捕捞等人类活动的影响, 莱州湾海洋生境遭到破坏, 众多生物的栖息地出现破碎化现象, 部分海区呈现荒漠化趋势。与其他海洋生物一样, 三疣梭子蟹的栖息地也在不断减少, 资源量持续衰退(卢晓等, 2018)。栖息地适宜性评价在内陆河流及湖泊生物资源中运用较为广泛, 在海洋中的运用起步较晚(龚彩霞等, 2011), 近年来研究集中在大洋鱼类生物资源, 对于近海生物资源有较大的应用空间, 国内有关三疣梭子蟹栖息地适宜性的研究尚未见报道, 亟待开展相关研究。
HSI模型是进行栖息地适宜性定量评价的经典方法, 该方法最早由美国地理调查局国家湿地研究中心鱼类与野生生物署于20世纪80年代初提出(U.S. Fish and Wildlife Service, 1981)。GAM模型常用来分析生物丰度和环境变量的非线性关系(Wood et al, 2002), 可以在HSI模型中选择环境变量。BRT模型基于决策树理论的学习方法, 可有效计算出变量因子对模型建立的贡献大小(De’ath, 2007), 生成多重回归树, 获取变量的权重分布。
本文构建了四种HSI模型, 包括普通HSI模型; GAM优化HSI模型; BRT优化HSI模型; GAM和BRT优化HSI模型, 对莱州湾三疣梭子蟹栖息地适宜性进行研究, 以期摸清资源分布状况, 明确适宜的环境特征, 全面了解其栖息地生存质量, 为三疣梭子蟹栖息地保护和莱州湾生态修复提供帮助。
1 材料与方法 1.1 数据来源和预处理三疣梭子蟹数据来源于2010~2020年夏季底拖网调查, 调查区域为莱州湾海域(119°00′~120°15′E, 37°12′~37°40′N), 调查站位每年20个(图 1)。调查船只功率260 kW, 调查网具为单船底拖网, 网口周长30.6 m, 囊网网目20 mm, 拖曳时网口宽度约8 m。
 |
| 图 1 莱州湾三疣梭子蟹调查站位 Fig. 1 Sampling stations of P. trituberculatus in the Laizhou Bay |
环境数据利用美国ORI-ON520M-01A便携式YSI水质分析仪现场测定, 内容为水深(depth, D)、底层水温(bottom sea temperature, BST)、底层叶绿素(bottom sea chlorophyll, BSC)、底层盐度(bottom sea salinity, BSS)。大型底栖生物(Macrozoobenthos, Mzb)取样使用开口面积为0.05 m2箱式采泥器采集, 每个站位取2个平行样, 使用0.05 mm的网筛分选大型底栖动物, 样品的保存、处理、计数和称量等均按《海洋调查规范》GB/T 12763.6—2007 (中华人民共和国国家质量监督检验检疫总局等, 2008)进行。
以调查网具拖速3 kn、拖网时间1 h为基准, 根据各站位实际拖网时间对调查数据进行标准化处理, 将其换算为单位时间的生物量(biomass, Bio) (g/h)和个数(individual, Ind) (ind./h), 以表征三疣梭子蟹的资源丰度。
1.2 研究方法 1.2.1 变量因子的选择GAM是一种非参数化的多元线性回归模型, 可以用来表示响应变量和解释变量的非线性关系, 本文利用GAM模型选择HSI模型中的变量。GAM模型通常表示为:
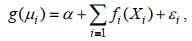 (1)
(1)式中, g为链接函数, μi =E(Yi), Xi为第i个响应变量的解释变量, β为模型估计参数, Yi为第i个响应变量, fi为平滑函数。将变量因子逐渐加入GAM模型中, 选取AIC值最小的模型作为最佳模型(Damalas et al, 2007)。AIC值的计算(Reid, 2003)如下:
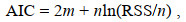 (2)
(2)式中, m为模型中参数的个数, n为观察值(数据样本)的个数, RSS为残差平方和。
1.2.2 变量因子权重分析BRT模型基于变量因子的相对贡献确定其在HSI模型中的权重(Chang et al, 2010; Yi et al, 2016), BRT方法是在运算过程中随机抽取一定量的数据, 分析自变量对因变量的影响程度, 而剩余数据用来对拟合结果进行检验, 对生成的多重回归树取平均值输出, 从而得出自变量对因变量的相对重要性(Keinath et al, 2017)。本文使用R 4.0.3中“GBM”包的“GBM.STEP”功能实现BRT模型, 其中抽样率(Train. Fraction)设置为0.8, 重复循环计算1 000次, 获取每个环境变量的相对贡献, 即环境变量的权重分布。
1.2.3 HSI模型首先, 利用样条平滑函数建立变量因子与适宜性指数(suitability index, SI)的关系模型, 分别以生物量Bio和个数Ind建立SI, 调查数据中Bio和Ind最小值为0, 三疣梭子蟹在调查海域资源密度不均, 最大值和最小值量级差距较大, 直接以Bio和Ind数据建模会降低SI指数精度, 本文对环境变量进行分组, 以各组距范围内Bio和Ind的总和建立每个组距下的SI。SI的取值范围为0~1, 一般认为0.7~1之间是适宜的栖息环境范围(Brooks, 1997; Xue et al, 2017), 计算公式如下:
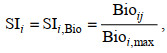 (3)
(3)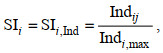 (4)
(4)式中, SIi为i环境变量的适宜性指数, SIi, Bio为以生物量Bio为基础建立的适宜性指数, SIi, Ind为以个数Ind为基础建立的适宜性指数, Bioij为i环境变量的j组距的Bio, Bioi, max为i环境变量的最大Bio, Indij为i环境变量的j组距的Ind, Indi, max为i环境变量的最大Ind。
HSI模型是由SI模型构建的综合模型, 取值范围为0~1, 代表栖息地适宜性“差”到“好”, 本文采用算数平均法(arithmetic mean model, AMM)和几何平均法(geometric mean model, GMM)来构建HSI模型, 计算公式分别为:
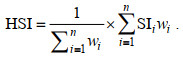 (5)
(5)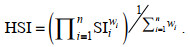 (6)
(6)采用2010~2018年数据构建HSI模型, 包括四种类型, 分别为: 普通HSI模型; GAM优化HSI模型(GAM对变量进行筛选后建立的HSI模型); BRT优化HSI模型(BRT对变量权重进行优化建立的HSI模型); GAM和BRT优化HSI模型。通过Pearson检验2020年实测值HSI和模型预测值HSI的相关性, 采用ArcGIS 10.2普通Kriging插值法绘制2020年莱州湾三疣梭子蟹HSI地图。
2 结果 2.1 变量因子的选择和权重分析以生物量表征资源丰度的GAM模型(表 1), 依次选取了D、BST、Mzb密度和BSS组成AIC值最小的模型, 累计解释方差为49.3%, D和BST与生物量极显著相关(P < 0.01), Mzb密度和BSS与生物量显著相关(P < 0.05)。以尾数表征资源丰度的GAM模型, 依次选取了D、BST、BSS和Mzb密度组成AIC值最小的模型, 累计解释方差为43.5%, D、BST和BSS与生物量极显著相关(P < 0.01), Mzb密度与生物量显著相关(P < 0.05)。
| 模型 | P值 | AIC | 累计解释方差/% |
| 生物量 | |||
| 水深 | 0.001 | 689.77 | 20.3 |
| 底层水温 | 0.001 | 687.52 | 38.4 |
| 大型底栖生物密度 | 0.032 | 686.55 | 45.9 |
| 底层盐度 | 0.035 | 685.32 | 49.3 |
| 尾数 | |||
| 水深 | 0.002 | 708.83 | 18.8 |
| 底层水温 | 0.002 | 707.76 | 34.2 |
| 底层盐度 | 0.006 | 706.50 | 39.8 |
| 大型底栖生物密度 | 0.044 | 705.50 | 43.5 |
由图 2可知, 以生物量和尾数表征资源密度构建的BRT模型变量因子的相对贡献顺序相同, BSS的相对贡献最大, 分别为26.9% (生物量)和23.5% (尾数), BSC的相对贡献最小, 分别为12.2% (生物量)和11.4% (尾数), 其他因子的相对贡献大小依次为BST、D和Mzb密度, 区间范围为19.0%~23.3%。
 |
| 图 2 以生物量和尾数表征资源密度构建的提升回归树模型 Fig. 2 P. trituberculatus abundance shown in the BRT model constructed in biomass and mantissa |
从变量因子的适宜性指数曲线(图 3)可以看出, 以生物量和尾数表征资源丰度拟合的三疣梭子蟹适宜栖息环境范围(SI > 0.7)基本相同。BST的适宜范围为27.2~28.5 ℃, BSS的适宜范围为25.5~30.5, D的适宜范围为9.0~10.5 m, Mzb密度的适宜范围为2 900~3 100个/m2, BSC浓度的适宜范围为13.5~ 14.5 mg/m3。
 |
| 图 3 以生物量和尾数表征资源丰度与环境因子的适宜性指数曲线 Fig. 3 Fitted suitability index (SI) curves based on the relationship between P. trituberculatus abundance in terms of biomass and mantissa and environmental factors |
以生物量表征资源丰度的GAM和BRT优化算术平均模型, 与2019~2020实际调查的生物量数据Pearson相关系数最高, 为0.653, 表明预测值与实测值有较强的相关性, 最优模型具有较强的预测能力。总体看来, 优化后的HSI模型均好于普通HSI模型, BRT优化的模型略好于GAM优化的模型, GAM和BRT优化的模型好于其他3种模型(表 2)。此外, 生物量表征资源丰度的模型好于尾数表征资源丰度的模型, 算术平均模型的相关系数高于几何平均模型。
| 资源丰度依据 | HSI模型 | 相关系数 | |
| 生物量 | 算术平均模型 | 普通 | 0.405 |
| BRT优化 | 0.551 | ||
| GAM优化 | 0.514 | ||
| GAM和BRT优化 | 0.653 | ||
| 几何平均模型 | 普通 | 0.341 | |
| BRT优化 | 0.504 | ||
| GAM优化 | 0.458 | ||
| GAM和BRT优化 | 0.536 | ||
| 尾数 | 算术平均模型 | 普通 | 0.378 |
| BRT优化 | 0.533 | ||
| GAM优化 | 0.512 | ||
| GAM和BRT优化 | 0.631 | ||
| 几何平均模型 | 普通 | 0.356 | |
| BRT优化 | 0.52 | ||
| GAM优化 | 0.496 | ||
| GAM和BRT优化 | 0.622 | ||
根据GAM和BRT优化的算术平均HSI模型预测结果可以看出, 伏季休渔结束前三疣梭子蟹主要分布在莱州湾南部、东南部和东北部海域, 中部和西南部海域分布较少, HSI值较低(图 4)。模型预测结果与2020年实际调查的三疣梭子蟹分布情况大体相符, 以生物量表征资源丰度的HSI模型预测的三疣梭子蟹最适海域(HSI > 0.7)范围明显大于以尾数表征资源丰度的HSI模型, 以生物量表征资源丰度的HSI模型对资源丰度较高区域的预测更加准确。
 |
| 图 4 2020年莱州湾三疣梭子蟹栖息地优化模型预测结果与实际调查情况分布图 Fig. 4 Spatial distribution of observed abundance of P. trituberculatus in 2020 overlaid with the predicted HSI values based on optimized HSI models |
生物的栖息地受多种因子的影响, 不同因子的影响大小不同, 合理选择影响因子和设置各因子的权重是优化HSI模型的两种主要方法。本研究分别采用GAM模型对三疣梭子蟹影响因子进行选择和BRT模型设置因子权重, 结果(表 2)显示BRT优化模型好于GAM优化模型, 说明在对HSI模型优化的过程中, 权重设置比因子选择更加重要, 与Zhang等(2020)研究结果一致。通过查阅文献资料基本可以囊括研究对象栖息地的大部分影响因子, 加上专家知识的判断筛选重要因子, 可以比较准确的完成因子选择的过程(龚彩霞等, 2011), 但权重设置受不同研究海域和不同研究对象的影响较大, 专家无法获得足够的信息对不同因子权重准确设置(Tian et al, 2009), 本研究利用BRT模型设置因子权重, 与非优化模型相比更具科学性和针对性, 将因子影响程度大小定量表示, 这也是导致权重设置比因子选择优化效果更好的原因。但仍不能忽视GAM模型对因子选择的重要性, 综合利用GAM模型和BRT模型才能构建最佳的HSI模型。
研究对象资源丰度指标的选取也是影响HSI模型建模效果的重要因素, 主要的资源丰度指标包括渔获量(范秀梅等, 2020)、渔获尾数、作业次数、单位捕捞努力量渔获量(catch per unit effort, CPUE) (刘瑜等, 2021)、生物量和渔获密度(朱承之等, 2020)等, 相关研究大都采用单一指标描述资源丰度, 本文比较了生物量和尾数表征三疣梭子蟹资源丰度对构建HSI模型的影响, 发现以生物量表征资源丰度的模型好于尾数, 在GAM模型的累计解释率也相对较高, 可能的原因是三疣梭子蟹个体大小差距较大, 部分站位大型个体较多, 用尾数表征降低了该区域的资源丰度, 但在适宜性指数曲线拟合得到的适宜栖息环境范围基本相同。范秀梅等(2020)分别基于渔获量和作业次数表征日本鲭鱼的资源丰度建立HSI模型, 发现两种方法在预报渔场上精度相差不大, 原因是日本鲭渔船作业次数和渔获量通常成正比关系, 作业次数反映了渔船的集中程度和渔民对产量的满意度(Bordalo-Machado, 2006), 作业次数多表明该海域有很高的渔获量。Tian等(2009)比较了基于CPUE和作业次数建立的西北太平洋柔鱼HSI模型, 结果表明, 基于CPUE的HSI模型高估了柔鱼的最佳栖息地范围, 基于作业次数的HSI模型能更好地定义柔鱼的栖息地范围。因此, 在选取资源丰度指标时, 应充分考虑研究对象的生物学以及作业方式等特性, 并采用多种指标表征资源丰度, 以获得更好的HSI模型。此外, HSI模型的计算方法中, 算术平均法AMM普遍具有较高的效果(陈新军等, 2009), 本文研究结果也证明了AMM的优势, 但相关研究表明GMM对SI的极大值和极小值较敏感(Vinagre et al, 2006)。
3.2 三疣梭子蟹栖息地的影响因素莱州湾三疣梭子蟹冬季在渤海10~30 m水深较深海域越冬, 4月开始生殖洄游, 到3~5 m深的港湾或河口等浅海附近海域产卵, 8月大多数三疣梭子蟹度过产卵期, 进入索饵育肥期, 是三疣梭子蟹资源量最高的时期, 9月伏季休渔结束后, 高强度的捕捞会使资源量迅速下降, 人为因素成为主导三疣梭子蟹栖息地分布的决定因素, 环境因素影响力下降(Tian et al, 2009), 因此8月伏季休渔结束前是研究三疣梭子蟹栖息地适宜性的最佳时期。本文研究了影响莱州湾三疣梭子蟹的部分环境因子和生物因子, GAM模型和BRT模型结果均表明BST、BSS和D对其栖息地影响较大, 与吴强等(2016a)研究结果相似。三疣梭子蟹是广盐性蟹类, 但其BSS的最适范围为25.5~30.5 (图 3), 夏季雨水丰富, 莱州湾淡水入海量增加, 近海盐度变化成为影响三疣梭子蟹栖息地的最主要因素(图 2)。三疣梭子蟹最适栖息地的BST范围为27.2~28.5 ℃, 水温不仅是决定三疣梭子蟹的季节性洄游的主要因素, 夏季是索饵育肥的重要时期, 水温过高会降低其体内消化酶的活性, 影响摄食行为(廖永岩等, 2008), 进而对栖息地分布产生影响。莱州湾大部分区域水深小于10 m, 最大水深18 m, 水深较浅的近岸水域是三疣梭子蟹产卵栖息的重要场所, 水深的变化会引起水温、盐度和营养盐等的改变, 对栖息地分布产生影响, 本文研究发现伏季休渔结束前三疣梭子蟹在莱州湾的最适水深为9.0~10.5 m。相关研究表明三疣梭子蟹分布与底栖生物的关系密切(吴强等, 2016b), 本文研究认为Mzb密度为2 900~3 100个/m2时是三疣梭子蟹最适范围, 底栖生物是大型甲壳类的重要食物来源, 三疣梭子蟹与日本蟳、口虾蛄和中国明对虾之间存在较强的食物竞争关系(李凡等, 2021), 导致底栖生物密度成为限制其栖息地分布的影响因素, 夏季莱州湾饵料食物较为丰富, 底栖生物与水温和盐度的影响相比较小, 在食物缺乏季节或海域, 底栖生物的影响力将增强。
3.3 夏季莱州湾三疣梭子蟹栖息地空间分布结合实际调查数据以及HSI模型的预测结果, 夏季莱州湾整体表现为近岸海域比离岸海域更适合三疣梭子蟹栖息, 这与徐炳庆等(2021)的研究结果一致, 8~9月份伏季休渔结束前水温较高, 三疣梭子蟹尚未进入越冬洄游阶段, 集中分布于近岸海域。本研究调查船只为底拖网渔船, 无法在水深低于5 m海区设置站位, 如采用小型船只或其他作业方式对浅海三疣梭子蟹资源进行调查, 将使近岸资源好于离岸资源的趋势更加明显。2020年莱州湾西南部近岸海域在HSI模型预测中栖息地适宜性较差, 主要原因为7211站位和7214站位底层平均水温30 ℃, 底层平均盐度23.8, 均处于适宜性指数曲线SI较低的位置, 导致HSI值较小, 成为不适宜三疣梭子蟹栖息的海域。但在2020年的实际调查中, 7214站位生物量较高, 为2 196 g/h, 可能的原因是站位设置中环境数据是与三疣梭子蟹资源数据同步, 近岸海域温盐环境变化较大, 站位设置稀疏使得环境数据的获取出现偶然性, 在以后的调查研究中, 应当对近岸海域环境监测站位设置更加紧密, 以获取更加准确地数据, 提高栖息地预测的精确性。
4 结论本研究运用GAM模型和BRT模型在建模过程中对HSI模型进行了优化, 优化模型对适宜栖息地预测效果均好于未优化模型, 优化模型对莱州湾三疣梭子蟹栖息地适宜性有较强的预测能力。优化模型显示BST、BSS和D对三疣梭子蟹栖息地影响较大, 莱州湾南部、东南部和东北部是夏季三疣梭子蟹的适宜栖息海域。
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. 海洋调查规范第6部分: 海洋生物调查: GB/T 12763.6-2007 [S]. 北京: 中国标准出版社: 1-157.
|
邓文洪, 2009. 栖息地破碎化与鸟类生存. 生态学报, 29(6): 3181-3187 DOI:10.3321/j.issn:1000-0933.2009.06.049 |
卢晓, 董天威, 涂忠, 等, 2018. 山东省三疣梭子蟹增殖放流回顾与思考. 渔业信息与战略, 33(2): 104-108 |
朱承之, 张云雷, 赛可, 等, 2020. 生物和非生物因子对秋季海州湾长蛇鲻栖息地适宜性的影响. 海洋学报, 42(6): 44-51 |
刘瑜, 花传祥, 2021. 基于GAM和权重分析的西北太平洋秋刀鱼渔情预报研究. 中国水产科学, 28(7): 888-895 |
李凡, 丛旭日, 张孝民, 2021. 莱州湾4种大型甲壳类的空间与营养生态位. 水产学报, 45(8): 1384-1394 |
吴强, 王俊, 陈瑞盛, 等, 2016a. 莱州湾三疣梭子蟹的生物学特征、时空分布及环境因子的影响. 应用生态学报, 27(6): 1993-2001 |
吴强, 金显仕, 栾青杉, 等, 2016b. 基于饵料及敌害生物的莱州湾中国对虾(Fenneropenaeus chinensis)与三疣梭子蟹(Portunus trituberculatus)增殖基础分析. 渔业科学进展, 37(2): 1-9 |
邹易阳, 薛莹, 麻秋云, 等, 2016. 应用栖息地适宜性指数研究海州湾小黄鱼的空间分布特征. 中国海洋大学学报, 46(8): 54-63 |
陈新军, 刘必林, 田思泉, 等, 2009. 利用基于表温因子的栖息地模型预测西北太平洋柔鱼(Ommastrephes bartramii)渔场. 海洋与湖沼, 40(6): 707-713 DOI:10.3321/j.issn:0029-814X.2009.06.006 |
范秀梅, 唐峰华, 崔雪森, 等, 2020. 基于栖息地指数的西北太平洋日本鲭渔情预报模型构建. 海洋学报, 42(12): 34-43 |
林群, 王俊, 李忠义, 等, 2015. 黄河口邻近海域生态系统能量流动与三疣梭子蟹增殖容量估算. 应用生态学报, 26(11): 3523-3531 |
易雨君, 程曦, 周静, 2013. 栖息地适宜度评价方法研究进展. 生态环境学报, 22(5): 887-893 DOI:10.3969/j.issn.1674-5906.2013.05.027 |
徐炜, 马志远, 井新, 等, 2016. 生物多样性与生态系统多功能性: 进展与展望. 生物多样性, 24(1): 55-71 |
徐炳庆, 陈玮, 王田田, 等, 2021. 莱州湾"伏休"结束前三疣梭子蟹的资源状况及其分布特征. 水产学报, 45(4): 543-551 |
龚彩霞, 陈新军, 高峰, 等, 2011. 栖息地适宜性指数在渔业科学中的应用进展. 上海海洋大学学报, 20(2): 260-269 |
廖永岩, 肖展鹏, 袁耀阳, 2008. 三疣梭子蟹幼体和幼蟹的温度适应性. 水生生物学报, 32(4): 534-543 DOI:10.3321/j.issn:1000-3207.2008.04.014 |
BORDALO-MACHADO P, 2006. Fishing effort analysis and its potential to evaluate stock size. Reviews in Fisheries Science, 14(4): 369-393 DOI:10.1080/10641260600893766 |
BROOKS R P, 1997. Improving habitat suitability index models. Wildlife Society Bulletin, 25(1): 163-167 |
CHANG J H, CHEN Y, HOLLAND D, et al, 2010. Estimating spatial distribution of American lobster Homarus americanus using habitat variables. Marine Ecology Progress Series, 420: 145-156 DOI:10.3354/meps08849 |
DAMALAS D, MEGALOFONOU P, APOSTOLOPOULOU M, 2007. Environmental, spatial, temporal and operational effects on swordfish (Xiphias gladius) catch rates of eastern Mediterranean Sea longline fisheries. Fisheries Research, 84(2): 233-246 DOI:10.1016/j.fishres.2006.11.001 |
DE'ATH G, 2007. Boosted trees for ecological modeling and prediction. Ecology, 88(1): 243-251 DOI:10.1890/0012-9658(2007)88[243:BTFEMA]2.0.CO;2 |
KEINATH D A, DOAK D F, HODGES K E, et al, 2017. A global analysis of traits predicting species sensitivity to habitat fragmentation. Global Ecology and Biogeography, 26(1): 115-127 DOI:10.1111/geb.12509 |
LIU S, SUN J S, HURTADO L A, 2013. Genetic differentiation of Portunus trituberculatus, the world's largest crab fishery, among its three main fishing areas. Fisheries Research, 148: 38-46 DOI:10.1016/j.fishres.2013.08.003 |
REID J R W, 2003. Experimental design and data analysis for biologists. Austral Ecology, 28(5): 588-589 DOI:10.1046/j.1442-9993.2003.01318.x |
TIAN S Q, CHEN X J, CHEN Y, et al, 2009. Evaluating habitat suitability indices derived from CPUE and fishing effort data for Ommatrephes bratramii in the northwestern Pacific Ocean. Fisheries Research, 95(2/3): 181-188 |
U. S. Fish and Wildlife Service, 1981. Standards for the development of habitat suitability index models [R]. Washington: Division of Ecological Services: 1-81.
|
VINAGRE C, FONSECA V, CABRAL H, et al, 2006. Habitat suitability index models for the juvenile soles, Solea solea and Solea senegalensis, in the Tagus estuary: Defining variables for species management. Fisheries Research, 82(1/2/3): 140-149 |
WOOD S N, AUGUSTIN N H, 2002. GAMs with integrated model selection using penalized regression splines and applications to environmental modelling. Ecological Modelling, 157(2/3): 157-177 |
XUE Y, GUAN L S, TANAKA K, et al, 2017. Evaluating effects of rescaling and weighting data on habitat suitability modeling. Fisheries Research, 188: 84-94 DOI:10.1016/j.fishres.2016.12.001 |
YI Y J, SUN J, ZHANG S H, 2016. A habitat suitability model for Chinese sturgeon determined using the generalized additive method. Journal of Hydrology, 534: 11-18 DOI:10.1016/j.jhydrol.2015.12.055 |
ZHANG Y L, YU H M, YU H Q, et al, 2020. Optimization of environmental variables in habitat suitability modeling for mantis shrimp Oratosquilla oratoria in the Haizhou Bay and adjacent waters. Acta Oceanologica Sinica, 39(6): 36-47 DOI:10.1007/s13131-020-1546-8 |