 2016, Vol. 47
2016, Vol. 47中国海洋湖沼学会主办。
文章信息
- 郑静静, 刘建勇, 刘加慧, 张嘉晨, 袁瑞鹏. 2016.
- ZHENG Jing-Jing, LIU Jian-Yong, LIU Jia-Hui, ZHANG Jia-Chen, YUAN Rui-Peng. 2016.
- 凡纳滨对虾(Litopenaeus vannamei)不同生长阶段体重的遗传参数和育种值估
- GENETIC PARAMETERIZATION AND BREEDING VALUE ON BODY WEIGHT OF LITOPENAEUS VANNAMEI IN DIFFERENT GROWTH STAGES
- 海洋与湖沼, 47(5): 1005-1012
- Oceanologia et Limnologia Sinica, 47(5): 1005-1012.
- http://dx.doi.org/10.11693/hyhz20160400075
-
文章历史
- 收稿日期:2016-04-05
- 收修改稿日期:2016-06-24




2. 湛江市国兴水产科技有限公司 湛江 524025
2. Zhanjiang Guoxing Biotech Co., Ltd., Zhanjiang 524025, China
凡纳滨对虾(Litopenaeus vannamei)具有温度、盐度适应范围广, 生长速度快, 抗病性强等优点, 深受虾农的青睐, 近年来已成为我国重要的经济虾类之一(马春艳等, 2011)。进行凡纳滨对虾遗传改良, 培育出具有生长速度快、成活率高等性状的新品种, 是我国对虾养殖业可持续发展的重要保障。以数量遗传学为基础的选择育种技术, 是改良凡纳滨对虾种质的重要手段(栾生等, 2013)。国外从1989年开始, 对凡纳滨对虾开展了生长、存活和抗病等性状的遗传改良工作(De Donato et al, 2001, 2005, 2008; Argue et al, 2002; Gitterle et al, 2005a, b; Kanchanachai et al, 2011)。国内凡纳滨对虾的育种工作近年来也取得较大进展, 目前已培育出“中科1号”、“中兴1号”等四个对虾养殖新品系, 生长和抗逆性状获得一定程度的提高(陈锚等, 2008; 黄永春等, 2010)。
遗传参数和育种值的估计是选择育种的基础工作, 它们对了解特定性状的遗传效应有一定的影响。国内外已对凡纳滨对虾(L.vannamei)的体质量性状的遗传参数进行了估计, 栾生等(2013)利用平均信息约束极大似然法对凡纳滨对虾体重、存活性状进行遗传参数的估计, 结果表明凡纳滨对虾体重的遗传力范围为0.19—0.43; De Donato等(2001)发现4个品系凡纳滨对虾体质量的遗传力估计值为0.65。对于对虾不同生长阶段遗传力估计的研究报道较多, 例如Gitterle等(2005a)对430个全同胞家系的凡纳滨对虾在98d和161d的体质量遗传力进行估计, 估计值为(0.17±0.04)—(0.24±0.05); Kanchanacha等(2011)对16个全同胞家系的凡纳滨对虾在共同环境养殖77d和124d的体重等多个生长性状进行遗传参数的估计, 在124d时体重的遗传力±标准差的估计值为0.37±0.14。可见, 国内外对凡纳滨对虾体重的遗传参数值不尽相同, 不同的群体, 同一群体不同的生长阶段、采用不同的模型等因素均会对遗传力估计值产生影响。白俊艳等(2004)发现即使利用同一种方法, 不同的模型所得到的估计值也是不同的。因此, 在估计凡纳滨对虾遗传参数之前应先构建一个合适的统计分析模型, 对多种可能的模型进行比较, 从而筛选出一个最合适的模型是准确估计遗传参数的基础。鉴此, 本研究结合广东海洋大学正在实施的遗传育种项目, 采用单性状动物模型BLUP法, 对凡纳滨对虾60、105和150日龄体重的遗传参数进行估计, 并进行不同模型的比较, 寻找适合不同生长阶段体重性状的模型, 旨在为凡纳滨对虾选择育种提供理论依据和技术参数, 以达到提高育种效率的目的。
1 材料与方法 1.1 亲本来源所用的亲本材料分别由15个凡纳滨对虾群体组成:美国迈阿密群体; 美国科纳湾群体; 美国夏威夷群体; 2013年美国SIS群体; 2014年美国SIS群体; 泰国正大群体; 中兴1号; 科海1号; 东方中科1号; 海大抗热家系(HD); 粤海选育群体; 来源于乌石sis抗病一代(K1代)的乌石抗病一代; 来源于广东海洋大学育种中心的海洋大学家系A, 海洋大学家系B, 海洋大学家系C。每个群体挑选出个体强健、活力良好、性腺发育成熟的雌雄各10尾, 作为育种亲本进行家系育种工作。对所选出的亲虾均测量体重体长后, 进行左侧眼柄套环标记。
1.2 亲虾催熟与家系建立标记后的所有雄虾, 放在同一个雄虾池里进行统一管理催熟; 所有标记的雌虾切除右侧眼柄后放在同一个雌虾池中统一管理催熟。每天选取性腺发育成熟的亲虾, 采用人工授精技术, 通过巢式设计(♂1 : ♀2)定向交尾完成受精。7天之内构建了30个全同胞家系和18个半同胞家系。
1.3 幼体培育和中间暂养每个家系随机挑取1000尾无节幼体分别放入48个注水400L(总体积为500L)的玻璃纤维桶中继续培育至仔虾。期间的饵料主要有虾片、黑粒、微藻类、丰年虫等, 每天投喂4次, 根据对虾的发育情况对投喂量和投喂比例进行适宜调整。每天换水30%—50%, 温度控制在每天温度控制在30℃±2℃, 尽量保持每个家系的培育条件一致。
1.4 共同环境养殖与数据测量幼体生长至3—4cm, 在每个个体的第5与第6腹节处注射不同颜色组合的荧光物作为家系标记。待荧光标记稳定后, 每个家系随机选择100尾个体放在同一大池中进行共同环境养殖, 养殖密度调整为100尾/m2。当个体平均日龄达到60d、105d、150d时, 每个家系随机选取30尾利用电子天平进行体质量的测量(精确到0.01g), 记录每个个体的网箱、测量日期等信息。
1.5 统计动物模型根据动物模型理论, 结合对虾育种的实际情况, 组建了下列四种模型进行遗传参数和育种值的估计:
模型A: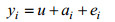
模型AF: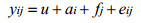
模型AD: 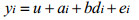
模型AFD: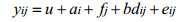
式中, u表示总体均值, yi, yij为第i个体体重, ai为第i个体育种值, fj为第j个家系的全同胞组效应, di、dij为测定体重的实际日龄协变量, b为回归系数, ei、eij为随机残差效应。
所用模型以矩阵形式表示为: y=Xb+Zu + e, 为观察值向量, b为固定效应向量, u为随机效应向量, e为随机误差向量, X为固定效应b的结构矩阵, Z为随机效应u的结构矩阵。数学期望定义为: E(u)=0, E(e)=0, E(y)=Xb; 方差定义为: 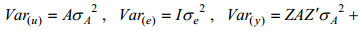
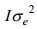
每个模型中包含的效应因子如表 1所示。
| 模型 | 随机效应 | 固定效应 | |
| 个体加性效应 | 全同胞组效应 | 日龄 | |
| A | √ | ||
| AF | √ | √ | |
| AD | √ | √ | |
| AFD | √ | √ | √ |
利用ASReml软件采用混合线性模型和约束极大似然法对凡纳滨对虾不同生长阶段体重性状的遗传参数和育种值进行估算。
1.7 不同动物模型的比较不同模型的比较通过似然比(LRT) (Kendall et al, 1979)进行检验, 检验公式为: 
AIC因能同时衡量模型的拟合度和精确度从而能对不同模型的优劣性进行判断, 对于一组模型, 可以认为AIC值最小的为最优模型(Akaike, 1976)。AIC=2K–2 lnL, K为参数个数, L为模型的最大似然函数值。
2 结果 2.1 4种模型估计不同生长阶段体重的方差组分及模型间的比较凡纳滨对虾在不同生长阶段体重的方差组分如表 2所示, 对于遗传力(h2)来说, 采用模型A所得到的遗传力估计值均比用其它模型得到的遗传力估计值大。全同胞组效应在各模型中的方差组分范围在0.06—0.12之间, 差别不大; 而各模型的残差组分差异较大, 60日龄的残差效应最大的是模型AFD, 为0.48, 最小的是模型AF, 为0.34; 105日龄的残差效应最大的是模型AFD, 为0.68, 最小的是模型A, 为0.31; 150日龄的残差效应最大的是模型AFD, 为0.72, 最小的是模型A, 为0.29。
| 日龄 | 模型 | σa2/σP2=h2(±SE) | σf2/σp2 | σe2/σP2 | 参数个数 | –2ln |
| 60日龄 | A | 0.64±0.07 | 0.36 | 2 | 5.40 | |
| AF | 0.54±0.08 | 0.12 | 0.34 | 3 | –9.79 | |
| AD | 0.62±0.06 | 0.38 | 2 | 5.49 | ||
| AFD | 0.39±0.07 | 0.13 | 0.48 | 3 | –9.51 | |
| 105日龄 | A | 0.69±0.05 | 0.31 | 2 | –9.97 | |
| AF | 0.48±0.04 | 0.08 | 0.44 | 3 | –8.87 | |
| AD | 0.37±0.08 | 0.63 | 2 | –11.10 | ||
| AFD | 0.25±0.04 | 0.07 | 0.68 | 3 | –10.54 | |
| 150日龄 | A | 0.71±0.07 | 0.29 | 2 | –12.39 | |
| AF | 0.54±0.15 | 0.06 | 0.40 | 3 | –12.36 | |
| AD | 0.31±0.05 | 0.69 | 2 | 5.49 | ||
| AFD | 0.22±0.05 | 0.06 | 0.72 | 3 | –12.30 | |
| 注: σp2为表型方差, σa2为加性遗传方差, σe2为残差, σf2为全同胞家系方差, h2为遗传力 | ||||||
凡纳滨对虾各生长阶段不同动物模型的似然比值和卡方检验的差异显著性如表 3所示, 在60日龄中, 不同模型之间的差异均极显著(P < 0.01); 105日龄不同模型之间的差异均不显著(P > 0.05); 150日龄, 模型A与模型AF、模型A与模型AFD之间差异不显著(P > 0.05), 模型AD与模型AFD之间的差异极其显著(P < 0.01)。凡纳滨对虾各生长阶段不同模型的AIC值见表 4, 当模型中考虑全同胞效应时, 可以看出模型AF和模型AFD的AIC值均比相对应的模型A和模型AD较为接近, 这说明将全同胞效应作为随机效应加入模型中是有必要的。当同时考虑日龄协变量时, 模型A与模型AFD比较仅在60日龄差异极显著(P < 0.01), 模型AD与模型AFD在60日龄、150日龄阶段差异均极显著(P < 0.01), 在105日龄阶段差异不显著(P > 0.05)。由于在模型中加入全同胞效应作为随机效应的优越性, 模型AD与模型AFD的比较更具代表性, AIC值的比较也说明模型AF与模型AFD相差不大:在60日龄, 模型AF的AIC值最小, 为-3.79, 比模型AFD小7.45%;在105日龄, 模型AFD的AIC值较小, 为-4.54, 比模型AF小36.78%;在150日龄, 模型AF的AIC值较小, 为-6.36, 比模型AFD小0.94%。考虑到模型AF估计的遗传力偏大和全同胞组效应确实存在的事实将模型AFD作为最优模型较为合适。遗传力的估计值在60、105和150日龄分别为0.39±0.07、0.25±0.04和0.22±0.05, 全同胞组效应分别为0.13、0.07和0.06。
| 模型比较 | 60日龄 | 105日龄 | 150日龄 |
| A : AF | 15.19** | –1.11 | –0.03 |
| A : AFD | 14.92** | 0.57 | –0.09 |
| AD : AFD | 15.00** | –0.55 | 17.79** |
| 注: *表示差异显著(P < 0.05), **表示差异极其显著(P < 0.01) | |||
| 模型 | 60日龄 | 105日龄 | 150日龄 |
| A | 9.40 | –5.97 | –8.39 |
| AF | –3.79 | –2.87 | –6.36 |
| AD | 9.49 | –7.10 | 9.49 |
| AFD | –3.51 | –4.54 | –6.30 |
分别利用家系育种值平均值和表型值平均值通过模型AFD对各家系不同生长阶段的家系育种值进行排序, 按照50%(前24位)的家系留取率, 两种选择方法的家系相同率分别为70.83%、62.50%、41.67%, 显然利用这两种选择方法选取的家系存在一定的差异(表 5)。60日龄、105日龄、150日龄利用育种值方法和表型值方法选取的前24个家系育种值平均值情况如表 5所示, 前者比后者分别高出约1.37%、19.97%、18.13%(图 1)。对家系进行选择时, 依据育种值选择比表型值选择更具优势。
| 日龄 | 家系相同率 (%) |
育种值平均值(g) | |
| 育种值比较 | 表型值比较 | ||
| 60日龄(AFD模型) | 70.83 | 1.48 | 1.46 |
| 105日龄(AFD模型) | 62.50 | 8.21 | 6.57 |
| 150日龄(AFD模型) | 41.67 | 13.21 | 11.19 |
 |
| 图 1 根据3个生长阶段的表型值排序和育种值排序分别选择前50%个家系的育种平均值比较 Fig. 1 The average breeding values of the first 50% of families selected in the sequence of phenotype and breeding value by optimal model |
分别利用育种值平均值和表型值平均值通过模型AFD对各家系不同生长阶段的个体育种值进行排序, 按照10%(前144个个体)的留种率, 三个生长阶段按照两种选择方法选取的个体相同率分别为60.08%、72.82%、65.53%, 这说明利用两种选择方法进行选取的个体存在一定的差异(表 6)。60日龄、105日龄、150日龄利用育种值方法和表型值方法选取的前144个个体育种值平均值情况如表 6所示, 前者比后者分别高出约20.06%、26.03%、12.01%(图 2)。对家系进行选择时, 依据育种值选择比表型值选择更具优势。
| 日龄 | 个体相同率(%) | 育种值平均值(g) | |
| 育种值比较 | 表型值比较 | ||
| 60日龄(AFD模型) | 60.08 | 3.19 | 2.55 |
| 105日龄(AFD模型) | 72.82 | 9.95 | 7.36 |
| 150日龄(AFD模型) | 65.53 | 13.07 | 11.50 |
 |
| 图 2 根据3个生长阶段的表型值排序和育种值排序分别选择前10%个体的育种平均值比较 Fig. 2 The average breeding values of the first 10% of individuals selected in the sequence of phenotype and breeding value by optimal model |
通过模型AFD计算每个生长阶段各家系和所有亲本(父本和母本)的的育种值, 均选取育种值排序前50%(前24位)的家系、亲本(即父本前12名, 母本前24名)进行比较。结果如表 7所示, 在60—105d、105—150d相邻生长阶段, 家系相同的个数分别为17、21, 家系的相同率分别为70.83%、87.50%;父本相同的个数分别为8、12, 对应的相同率分别为66.67%、100%;母本相同的个数分别为14、21, 对应的相同率分别为58.33%、87.50%;可见不同生长阶段根据家系、亲本育种值留取家系、亲本的差异较大, 随着生长期的增长, 根据体重育种值留取家系、亲本的差异越小。根据所有家系、亲本育种值的相关分析表明(表 7), 在60—105d、105—150d相邻生长阶段, 家系、父本、母本育种值相关系数的可信度均达到极显著水平(P < 0.01), 由上述结果可知, 不同生长阶段家系、父本和母本育种值相关性的变化趋势均为生长期越长, 相关性越高。
| 相邻生长阶段 | 家系 | 父本 | 母本 | |||||
| 相同率(%) | 相关系数 | 相同率(%) | 相关系数 | 相同率(%) | 相关系数 | |||
| 60—105日龄 | 70.83 | 0.589** | 66.67 | 0.511** | 58.33 | 0.458** | ||
| 105—150日龄 | 87.50 | 0.974** | 100 | 0.995** | 87.50 | 0.977** | ||
| 注: *表示差异显著(P < 0.05), **表示差异极显著(P < 0.01) | ||||||||
利用动物模型对不同的固定因素和随机因素进行多方面结合, 会得到不同的遗传参数和育种估计值, 因此, 应结合选择育种的实际情况, 对多种可能的模型进行比较分析, 从中找出一个最适合的模型(白俊艳等, 2004)。本研究对全同胞家系效应和日龄协变量进行考虑, 建立了4种模型对不同生长阶段体重的遗传参数进行了估计, 并利用似然比(LRT)及AIC值检验出最优模型计算育种值。由表 3可以看出, 当同时考虑日龄协变量时, 模型A与模型AFD的比较仅在60日龄差异极显著(P < 0.01), 模型AD与模型AFD在60日龄、150日龄阶段差异均极显著(P < 0.01), 在105日龄阶段差异不显著(P > 0.05)。而根据凡纳滨对虾各阶段不同模型的AIC值(表 4), 当仅考虑全同胞效应作为随机因子时, 模型AF和模型AFD的AIC值均比相对应的模型A和模型AD的AIC值较为接近, 这说明将全同胞效应加入模型中是有必要的。由于在模型中加入全同胞效应, 如表 3所示, 模型AD与模型AFD的比较更具代表性, 而从表 4结果可以看出模型AF和模型AFD的AIC值均比相对应的模型A和模型AD的AIC值较为接近, 但考虑到模型AF估计的遗传力偏大和全同胞组效应确实存在的事实, 因此用模型AFD对凡纳滨对虾不同生长阶段的遗传参数和育种值进行估计最为适合。
3.2 凡纳滨对虾不同生长阶段体重性状的遗传力估计遗传力是最重要的遗传参数之一, 通过对遗传力的估计, 可令下一代的遗传增益实现最大化。栾生等(2013)研究发现凡纳滨对虾体重的遗传力范围为0.19—0.43; De Donato等(2001)研究发现凡纳滨对虾体质量的遗传力估计值为0.65; Gitterle等(2005a)研究发现凡纳滨对虾在98日龄和161日龄的体质量遗传力估计值为(0.17±0.04)—(0.24±0.05); Kanchanacha等(2011)研究发现凡纳滨对虾在共同环境养殖124日龄时体重的遗传力±标准差的估计值为0.37±0.14;Castillo-Juárez等(2007)分别利用单性状线性混合模型与多变量线性混合模型评估凡纳滨对虾体质量的遗传力, 估计值分别为0.24—0.35与0.37—0.45。以上结果均表明凡纳滨对虾体质量遗传力均为中高等遗传力。本研究中, 由模型AFD估计的体重遗传力估计值在60、105和150日龄分别为0.39±0.07、0.25±0.04和0.22±0.05, 这与上述国内外的研究结果基本一致。本研究发现, 三个时间阶段的遗传力估计值分别为0.39±0.07、0.25±0.04和0.22±0.05, 遗传力值随着生长周期的增大而逐渐降低, 这可能是随着生长周期的增长, 凡纳滨对虾受母体效应影响逐渐减小的结果, 这与唐瞻杨等(2015)估计尼罗罗非鱼不同月龄体重、全长、体长等9个数量性状的遗传力结果相似。
3.3 动物模型BLUP方法估计凡纳滨对虾不同生长阶段体重性状育种值动物模型BLUP法因考虑了个体所有的血缘关系, 结合多方面的信息来源(如种群、世代、性别、池塘号、全同胞组等)对固定效应和随机因素进行深层次的剖分, 从而有助于提高育种值估计的准确性(盛志廉等, 2001; 栾生等, 2008)。在本研究中, 凡纳滨对虾生长处于60、105和150日龄3个阶段中, 利用育种值方法选取的前24个家系育种值平均值比利用表型值方法选取的分别高出约1.37%、19.97%、18.13%;利用育种值方法选取的前10%个个体育种值的平均值比利用表型值方法选取的分别高出约20.06%、26.03%、12.01%。由此可见, 利用动物模型进行选育与直接利用表型值选择相比较, 动物模型具有更高的优越性。众多研究表明, BLUP方法是目前估计育种值的最佳方法且获得显著的育种成效, 而动物模型BLUP方法的优势更为突出。Gall等(2002)研究发现, 利用动物模型BLUP法对罗非鱼体重性状进行选育, 每一代的平均遗传获得为(2.61±0.05)g, 进行三代选育后, 体重比基础群体增加了约40%, 而Hulata等(1986)和Huang等(1990)却发现采用群体选择法对罗非鱼进行选育, 得出的选育结果却为负值, 与Gall等(2002)利用动物模型BLUP法进行选育得出的结果形成鲜明的对比。众多水产动物研究结果也表明利用育种值选择, 选择效率明显高于表型值选择效率:如张天时等(2008)对145日龄的中国对虾分别利用育种值和表型值对留种家系和个体进行选择, 结果显示, 利用前者选择的家系或个体的育种值的平均值比利用后者选择分别高出50%和80.59%;罗坤等(2008)对混养88天的罗氏沼虾进行了体重育种值和表型值差异的比较, 结果发现家系育种值比表型值选择提高20.7%; 马爱军等(2009)对不同生长阶段大菱鲆的体重利用育种值选择和表型值选择两种方法分别对留种家系和个体进行选择, 结果表明在家系选择中, 前者比后者分别高出约3.1%、14.4%、20.5%、18.5%、24.8%, 在个体选择中, 前者比后者分别高出约12.15%、22.85%、14.15%、16.47%、12.78%。可见, 无论是对家系或者个体进行选择, 依据育种值选择比表型值选择表现得更具优势。
3.4 凡纳滨对虾家系筛选时期和亲本选择的时期选择育种是一项耗时长且投入大的工作, 我们在此过程中通过对不同生长阶段生长性状的遗传参数进行评估(特别是早期生长阶段和收获阶段生长性状的遗传相关), 可为准确评估多阶段选育的效果提供参考。如果早期生长阶段与收获阶段体重的遗传相关性很高, 对选育品种进行早期筛选可以减少养殖成本, 缩短繁殖周期等。国内外对凡纳滨对虾不同生长阶段的遗传参数估计工作开展得较多(De Donato et al, 2001, 2005, 2008; Argue et al, 2002; Gitterle et al, 2005a, b; 陈锚等, 2008; 黄永春等, 2010; Kanchanachai et al, 2011; 栾生等, 2013), 但是该何时对家系和亲本进行筛选的研究较少, 本文通过估算不同生长阶段的家系育种值及亲本育种值分别进行了家系和亲本的选择比较, 结果表明, 在60—105d、105—150d相邻生长阶段, 排名前50%的家系相同率分别为70.83%、87.50%, 父本相同率分别为66.67%、100%, 母本相同率分别为58.33%、87.50%, 可见不同生长阶段, 根据育种值留取家系和亲本的差别较大, 家系与亲本的变化趋势相同, 均为生长期越长, 差异越小。根据所有家系和亲本育种值的相关分析表明(表 7), 在60—105d、105—150d相邻生长阶段, 家系育种值的相关系数分别为0.589、0.974, 父本育种值的相关系数分别为0.511、0.995, 母本育种值的相关系数分别为0.458、0.977, 相关系数的可信度均达到极显著水平(P < 0.01), 由此可见60—105d日龄与105—150d日龄相比较, 生长期越长, 育种值的相关性越高。这是由于生物体在生长发育过程中, 其数量性状的基因表达受到时间(自身的生长发育期)和空间(生长周边环境)等外部条件的影响(Atchley et al, 1997), 生长期越短, 生长性能受内部和外界的影响越大, 表现得越不稳定。由于105—150d日龄较60—105d日龄3种相关性提高的幅度均显著增加, 因此可认为, 对家系及父本、母本进行提前筛选的时期至少在105日龄以上。可见凡纳滨对虾的早期生长阶段和收获阶段生长性状的遗传相关还需进一步研究。
4 结论本研究根据实际情况筛选出估计凡纳滨对虾体重性状的最优模型, 并对凡纳滨对虾不同生长阶段体重的遗传参数和育种值进行估计, 还对不同生长阶段的家系及亲本进行选择比较, 结果表明, 凡纳滨对虾的体重遗传力为中高度遗传力, 说明对其进行选择育种具有较大的潜力, 下一代可获得较大的遗传增益; 在不同生长阶段, 利用BLUP法育种值选择比表型值选择更具优势, 可以更加有效地提高凡纳滨对虾的遗传改良速度; 不同生长阶段家系、亲本育种值相关性的变化均为生长期越长, 相关性越高, 推测对家系及亲本进行提前筛选的时期至少在105日龄以上。
| 马春艳, 马洪雨, 马凌波, 等, 2011. 凡纳滨对虾引进群体和2个养殖群体遗传变异的微卫星分析. 海洋渔业, 33 (1) : 1–8 |
| 马爱军, 王新安, 雷霁霖, 等, 2009. 大菱鲆(Scophthalmus maximus)不同生长阶段体重的遗传参数和育种值估计. 海洋与湖沼, 40 (2) : 187–194 |
| 白俊艳, 李金泉, 道尔吉, 等, 2004. 用不同模型估计绒山羊早期生长性状遗传参数的比较. 遗传学报, 31 (6) : 578–581 |
| 张天时, 栾生, 孔杰, 等, 2008. 中国对虾体重育种值估计的动物模型分析. 海洋水产研究, 29 (3) : 7–13 |
| 陈锚, 吴长功, 相建海, 等, 2008. 凡纳滨对虾的选育与家系的建立. 海洋科学, 32 (11) : 5–8 |
| 罗坤, 孔杰, 栾生, 等, 2008. 应用动物模型对罗氏沼虾育种值估计的差别分析. 海洋水产研究, 29 (3) : 85–91 |
| 栾生, 孔杰, 王清印, 2008. 水产动物育种值估计方法及其应用的研究进展. 海洋水产研究, 29 (3) : 101–107 |
| 栾生, 罗坤, 阮晓红, 等, 2013. 凡纳滨对虾(Litopenaeus vannamei)体重、存活性状的遗传参数和基因型与环境互作效应. 海洋与湖沼, 44 (2) : 445–452 |
| 唐瞻杨, 陈文治, 罗永巨, 等, 2015. 尼罗罗非鱼(Tilapia nilotica)不同月龄数量性状遗传力估计. 海洋与湖沼, 46 (5) : 1180–1185 |
| 黄永春, 艾华水, 殷志新, 等, 2010. 第四代凡纳滨对虾抗WSSV选育家系的抗病及免疫特性研究. 水产学报, 34 (10) : 1549–1558 |
| 盛志廉, 陈瑶生, 2001. 数量遗传学. 北京: 科学出版社, |
| Akaike H, 1976. An information criterion (AIC). Mathematical Science, 14 (153) : 5–9 |
| Argue B J, Arce S M, Lotz J M, et al, 2002. Selective breeding of Pacific white shrimp (Litopenaeus vannamei) for growth and resistance to Taura Syndrome Virus. Aquaculture, 204 (3-4) : 447–460 DOI:10.1016/S0044-8486(01)00830-4 |
| Atchley W R, Zhu J, 1997. Developmental quantitative genetics, conditional epigenetic variability and growth in mice. Genetics, 147 : 765–776 |
| Castillo-Juárez H, Casares J C Q, Campos-Montes G, et al, 2007. Heritability for body weight at harvest size in the Pacific white shrimp, Penaeus (Litopenaeus) vannamei, from a multi-environment experiment using univariate and multivariate animal models. Aquaculture, 273 (1) : 42–49 DOI:10.1016/j.aquaculture.2007.09.023 |
| De Donato M, Cabrera S, Ramirez R et al, 2001. Analysis of growth in families of Litopenaeus vannamei under culture conditions in Venezuela. In: Aquaculture: Book of Abstracts. Baton Rouge, Louisiana: World Aquaculture Society, 168 |
| De Donato M, Manrique R, Ramirez R, et al, 2005. Mass selection and inbreeding effects on a cultivated strain of Penaeus (Litopenaeus) vannamei in Venezuela. Aquaculture, 247 (1-4) : 159–167 DOI:10.1016/j.aquaculture.2005.02.005 |
| De Donato M, Ramirez R, Howell C, et al, 2008. Artificial family selection based on growth rate in cultivated lines of Litopenaeus vannamei (Decapoda, Penaeidae) from Venezuela. Genetics and Molecular Biology, 31 (4) : 850–856 DOI:10.1590/S1415-47572008000500009 |
| Gall G A E, Bakar Y, 2002. Application of mixed-model techniques to fish breed improvement: analysis of breeding-value selection to increase 98-day body weight in tilapia. Aquaculture, 212 (1-4) : 93–113 DOI:10.1016/S0044-8486(02)00024-8 |
| Gitterle T, Rye M, Salte R, et al, 2005a. Genetic (co) variation in harvest body weight and survival in Penaeus (Litopenaeus) vannamei under standard commercial conditions. Aquaculture, 243 (1-4) : 83–92 DOI:10.1016/j.aquaculture.2004.10.015 |
| Gitterle T, Salte R, Gjerde B, et al, 2005b. Genetic (co) variation in resistance to White Spot Syndrome Virus (WSSV) and harvest weight in Penaeus (Litopenaeus) vannamei. Aquaculture, 246 (1-4) : 139–149 DOI:10.1016/j.aquaculture.2005.02.011 |
| Huang C M, Liao I C, 1990. Response to mass selection for growth rate in Oreochromis niloticus. Aquaculture, 85 (1-4) : 199–205 DOI:10.1016/0044-8486(90)90019-J |
| Hulata G, Wohlfarth G W, Halevy A, 1986. Mass selection for growth rate in the Nile tilapia (Oreochromis niloticus). Aquaculture, 57 (1-4) : 177–184 DOI:10.1016/0044-8486(86)90195-X |
| Kanchanachai Y, Poompuang S, Koonawootrittriron S, et al, 2011. Estimating genetic parameters for weight and body size of Pacific white shrimp (Litopenaeus vannamei) by restricted maximum likelihood method. Kasetsart Journal (Natural Sciences), 45 (6) : 1047–1057 |
| Kendall M, Stuart A, 1979. The Advanced Theory of Statistics. Vol.2. Inference and Relationship. London, UK: Charles Griffin & Company, |